Le noyau interphasique
On distingue les eucaryotes, des procaryotes en fonction du noyau. Ces derniers présentent différents aspects au cour de la vie cellulaire.
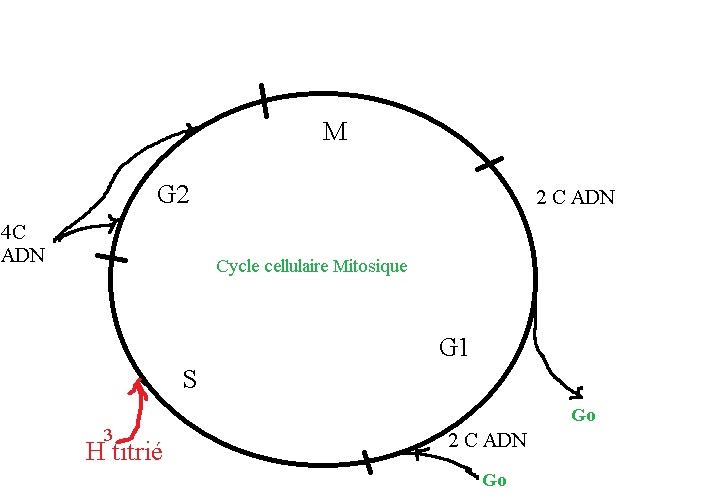
1/ Le cycle cellulaire:
État haploïde (23 chromosomes) à C = Quantité d'ADN
Pendant la mitose, le noyau a un aspect condensé, prenant fortement les colorants basiques.
A l'interphase, les chromosomes sont "décondensés" et forment la Chromatine, c'est la période la plus active avec:
-G1 La phase de transcription avec la synthèse d'ADN, ARN,...
-S Synthèse d'ADN
-G2 Phase pré-mitotique
- M Division cellulaire
Pour les cellules qui ne se divisent plus, il n'y a pas de phase S, elles vont en phase Go définitivement ou transitoirement. Par exemple: les neurones ou certaines qui sont en attentes de stimulations. Le marquage de l'ADN par la radioactivité (acide animé: thimidine H3 Titrié) se fait pendant la phase S.
2/ Structure générale du noyau observée en phase G1 et G2
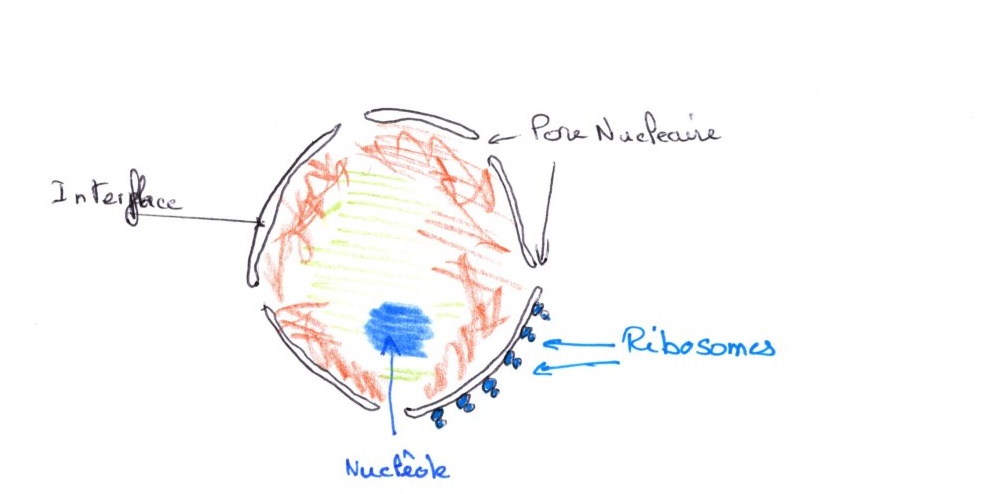
On distingue deux densités différentes:
-Une dense, très colorables, souvent fixée sur l'enveloppe, c'est l'hétérochromatine.
-Une plus claire, au centre, c'est l'euchromatine
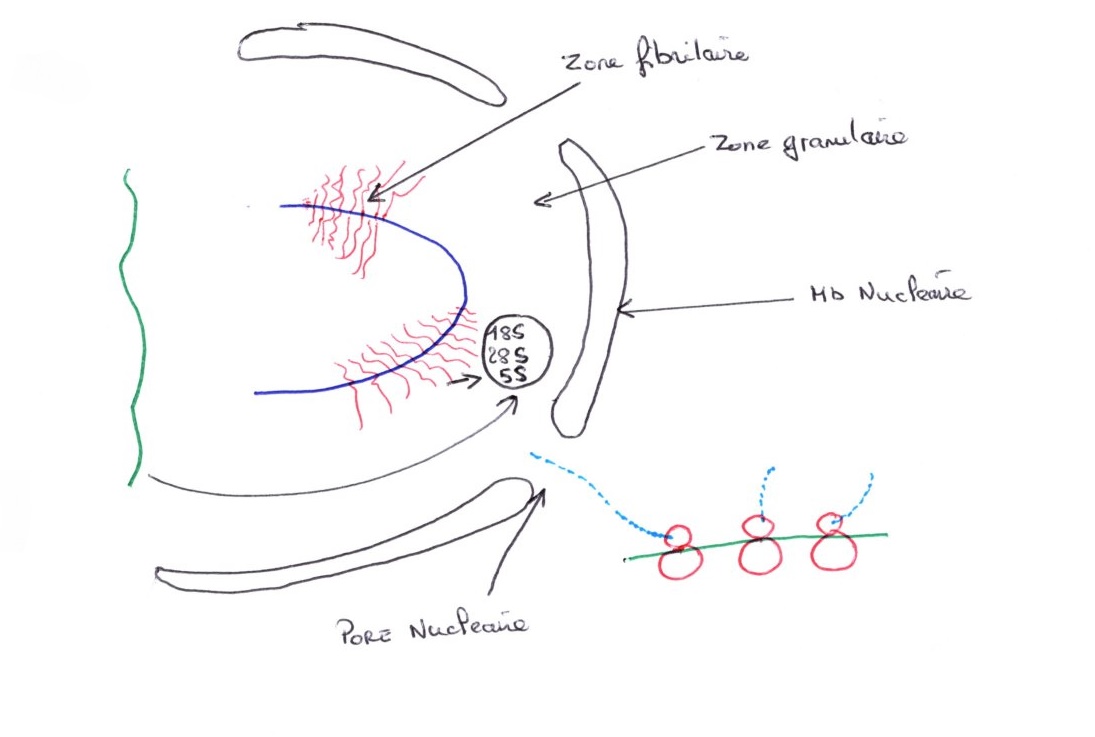
Le nucléole est très dense en microscopie électroniques et est riche en protéines plus ou moins grosses. Le nucléoplasme est la phase soluble qui imprègne le nucléole et les deux types de chromatine. Les pores nucléaires représentent le lieu de passage des macromolécules entre le noyau et le cytoplasme.
Observation au microscope électronique à haute résolution:
Le nucléole est très dense au microscope électronique, il est riche en protéines et est plus ou moins gros. le nucléoplasme est une phase soluble qui imprègne le nucléole et les deux types de chromatine. les ports nucléaires représentent le lieu de passage des macromolécules entre le noyau et le cytoplasme.
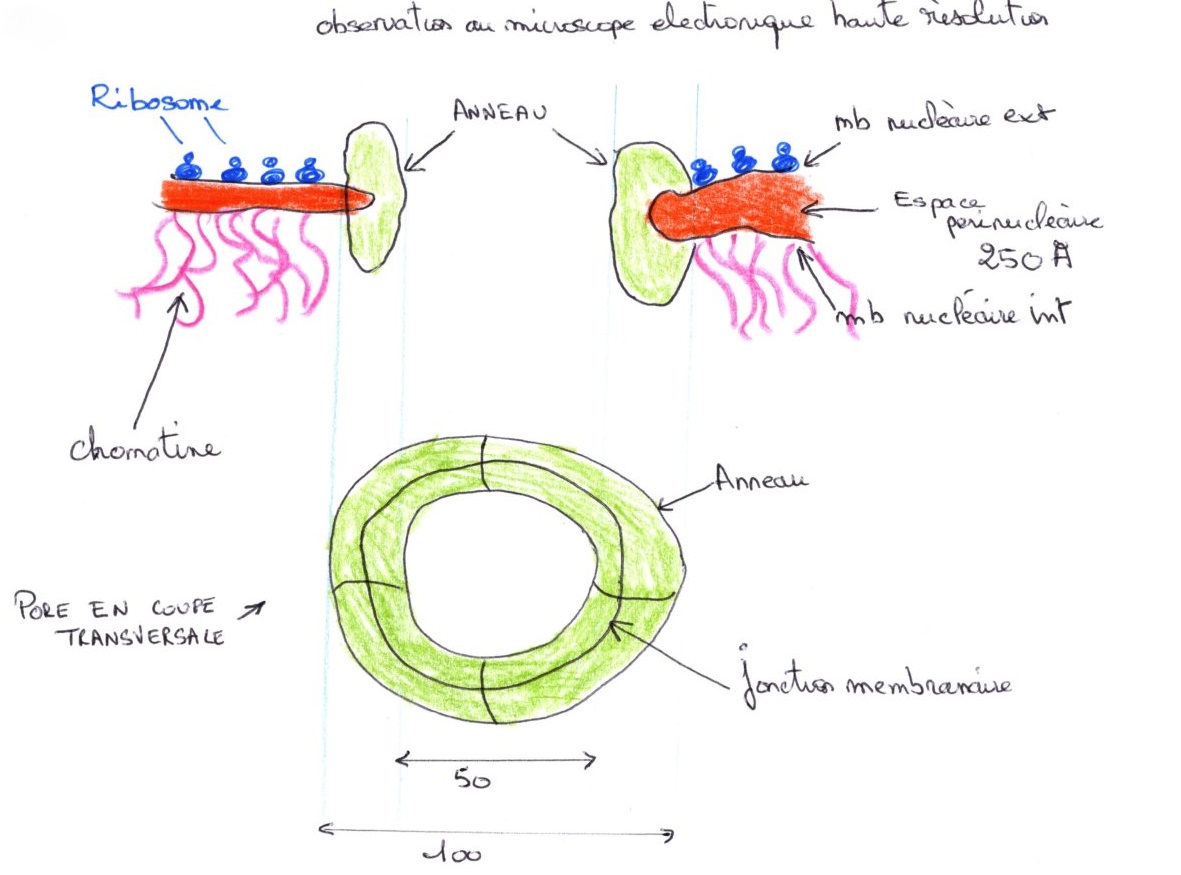
Le complexe du pore correspond à 8 sous-unités qui définissent un tunnel. La chromatine est fortement attachée à la lamina et elle ne flotte pas librement dans le nucléoplasme. Le nombre et la densité des pores sont variables d'une cellule à l'autre et est à peu près proportionnelle à l'activité de transcription.
Cas de l'ovocyte de xénope (amphibien):
Nature et composition des pores nucléaires :
digestion enzymatique, deux types de traitement :
- par des protéases : trypsine,...
- par des nucléases: DNases, RNases,...
Le traitement par des protéases ou les nucléases, ont le même effet, il y a disparition de l'anneau qui est digéré. Cela permet de dire qu'il est de nature Ribonucléoprotéique (RNP). Cette observation permet de penser, qu'il y a une association entre les protéines qui forment le complexe et les molécules qui traversent le pore. Lle passage par le port n'est pas un simple passage passif.
La membrane nucléaire est plus ou moins ressemblante à celle du RE, du point de vue pourcentage de protéines et de lipides. L'enveloppe nucléaire est en continuité avec le REG. Quand on localise des enzymes dans le REG, on peut aussi les retrouver dans l'espace périnucléaire, comme la cytochrome 450. C'est un compartiment de stockage, qui n'a pas les mêmes activités enzymatiques que le RE. La concentration en eau du nucléoplasme est inférieure à celle du cytoplasme.
2/Composition chimique :
L'ADN contient des caractères héréditaires et représente une constante de l'espèce point il s'agit d'une molécule longue, bicaténaire, fortement contrainte dans le noyau.
Mesure de la quantité d'ADN dans différentes espèces ( idée d'évolution des espèces avec la quantité d'ADN)
On s'aperçoit que la quantité d'ADN n'a pas de signification génétique pour l'espèce, ce n'est pas la quantité qui compte mais la qualité du contenu.
L'ARN du noyau est très hétérogène et est différent de ceux du cytoplasme. On distingue des précurseurs qui subissent une maturation avant d'aller dans le cytoplasme.
Dans le noyau on trouve :
- les pré-messagers qui sont de grosses molécules à courte durée de vie. 90 % d’entre elles sont dégradées en 10 à 30 minutes dans le noyau. ils ne passeront pas dans le cytoplasme, seuls les 10 % restants y passeront et se sont les ARN messager.
- les pré-ribosomes sont plus gros et seulement 10 % passeront dans le cytoplasme. Leur stockage et leur maturation ont lieu dans le nucléole.
- les ARN associés au nucléole et aux chromosomes, ont un poids moléculaire variant entre 4,5s et 10s. On ne les trouve pas dans le cytoplasme. Leur durée de vie est longue.
- les ARN initiateurs de la réplication de l'ADN sont très petits et sont dégradés après avoir joué leur rôle d'initiation. Leur présence empêche la fermeture des deux chaînes de l'ADN.
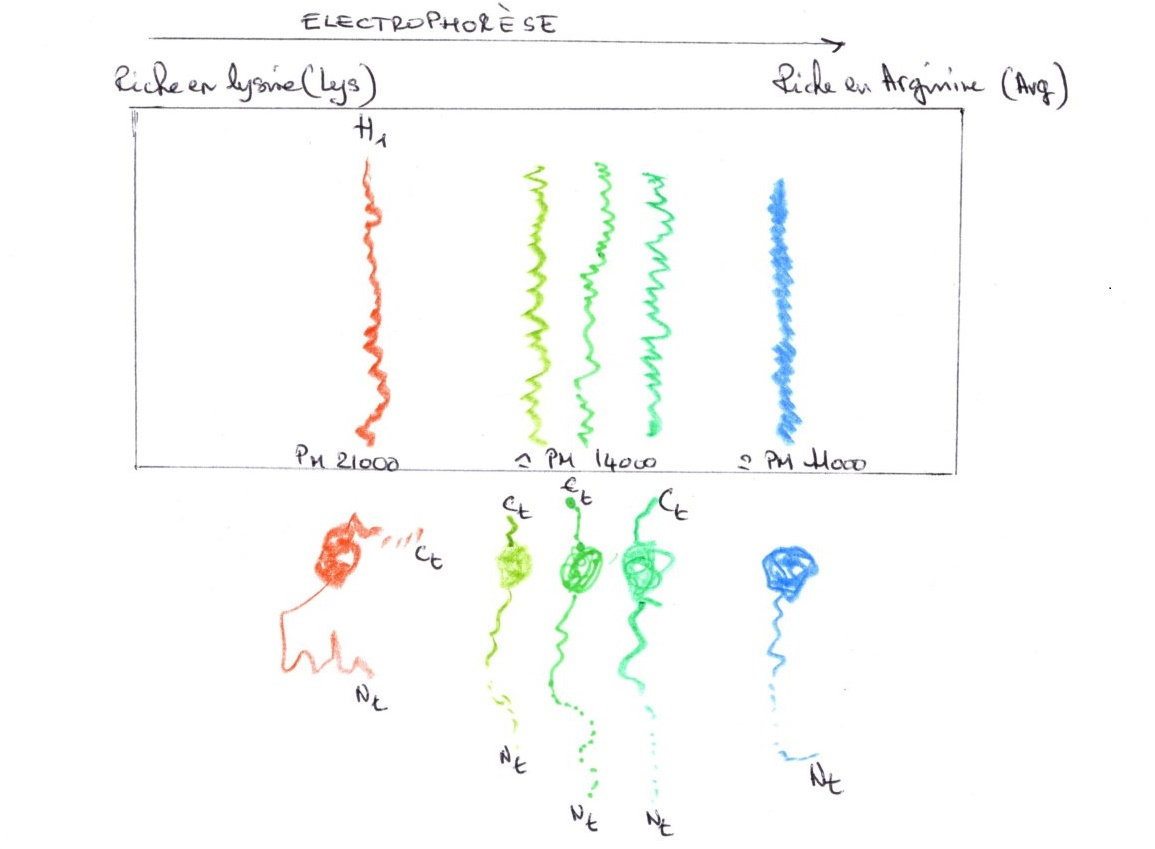

B/Grâce à l'hybridation un tri peut-être effectué parmi les gènes:
a-Séquences très courtes, répétées un grand nombre de fois, réunies en bloc dans la même région de la chromatine.
Exemple : chez la drosophile, l'ADN satellite est formé uniquement de deux types de bases. L'une des chaînes peut-être constituée uniquement de base puriques, l'autre de base pyrimidique. Cet ADN représente 10 à 25 % du génome total. Il est très faiblement transcrit. Il y a plus ou moins une inactivité génétique de l'ADN satellite, c'est de la chromatine peu active.
b-Gènes codés environ 10 fois, sont de deux types:
-gènes ribosomaux dans le nucléole, ils sont transcrits en ARN 45s mais non traduits.
-gènes codants pour les protéines de structure, il y a environ 10 gênes codants pour les histones, chez l'homme, la souris, l'amphibien xénope mais ils sont trop nombreux pour être purifiés.
Cas particulier : l'oursin pour lequel il existe 300 à 1000 copies par lot haploïde, on pourrait le considérer comme de l'ADN satellite, mais ces gènes sont transcrits très activement en protéines histones. C’est chez l'oursin que les gènes histones ont été observés, car on peut directement les utiliser comme sonde : l'ARN messager est utilisé comme sonde dans les cellules de l'oursin pour reconnaître le génome.
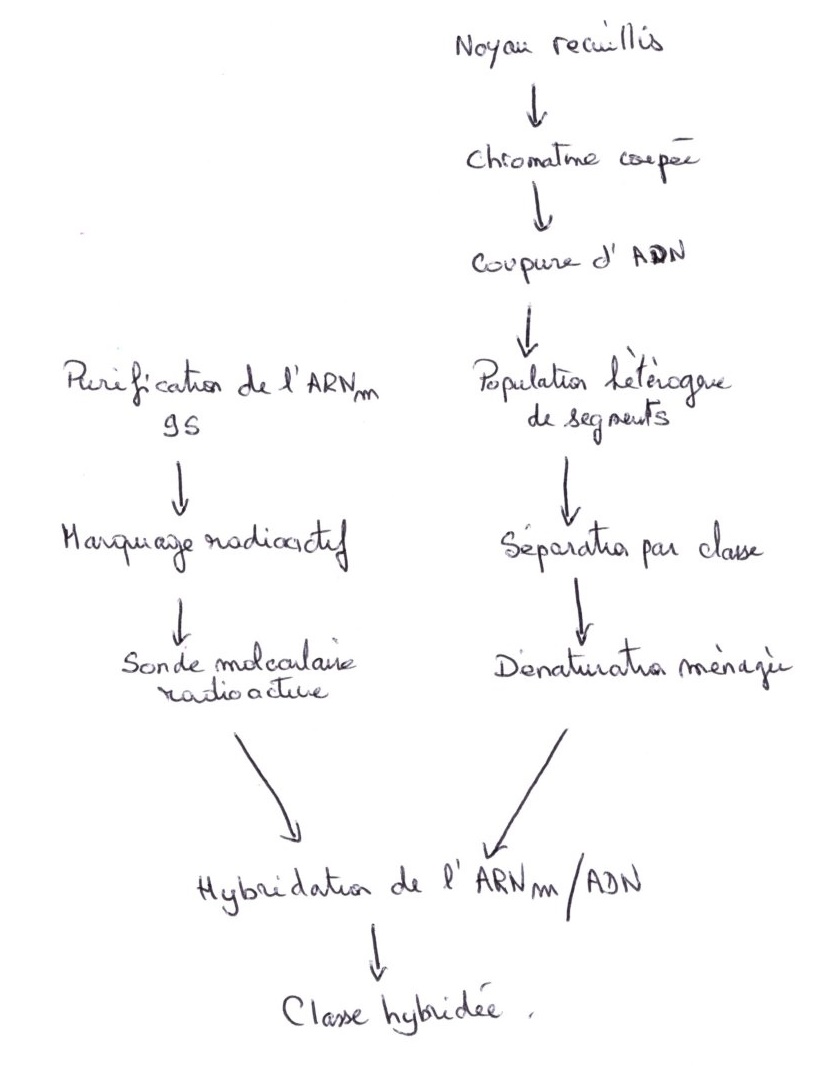
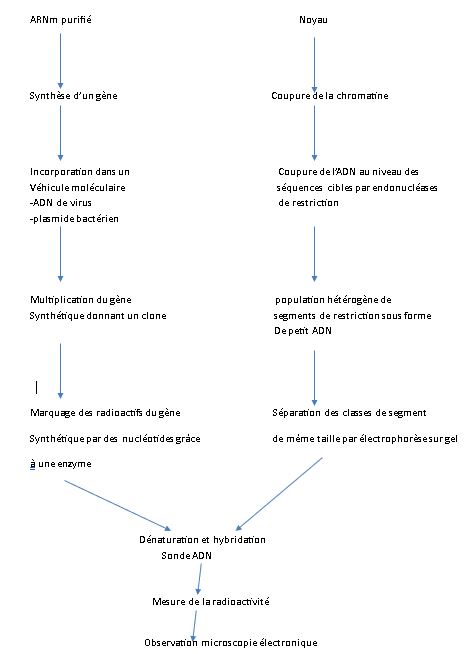
On purifie les ARNm messager 9S matures, traduits en histones dans le cytoplasme. Le marquage radioactif de cet ARNm fournit une sonde radioactive. Parallèlement, la chromatine du noyau est isolée. l'ADN est coupé en segment de restriction de taille inégale, puis séparé par classe de longueur identique. On effectue ensuite une dénaturation ménagée par chauffage modéré pour couper les liaisons hydrogène, puis on effectue une hybridation avec la sonde ARNm/ ADN.
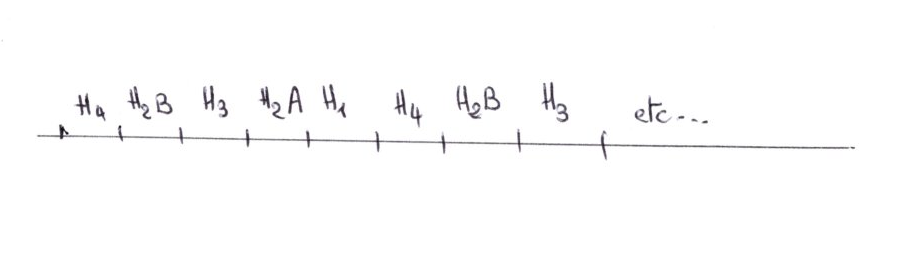
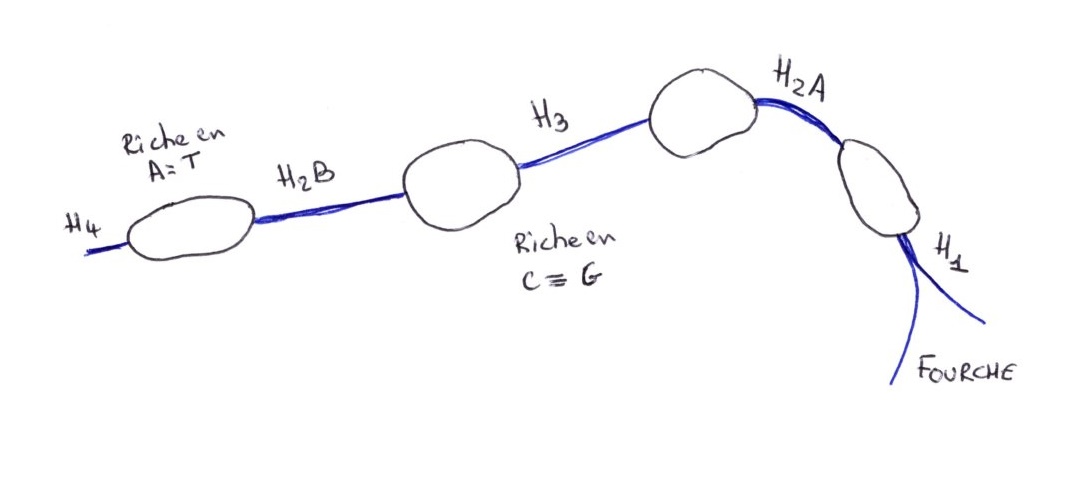
La classe radioactive est conservée, elle contient la sonde et permet une cartographie de chevauchement. L'ordre de succession des 5 gènes codants pour les 5 histones, et toujours la même.
H4; H2b;H3;H2n;H1;H4;.....
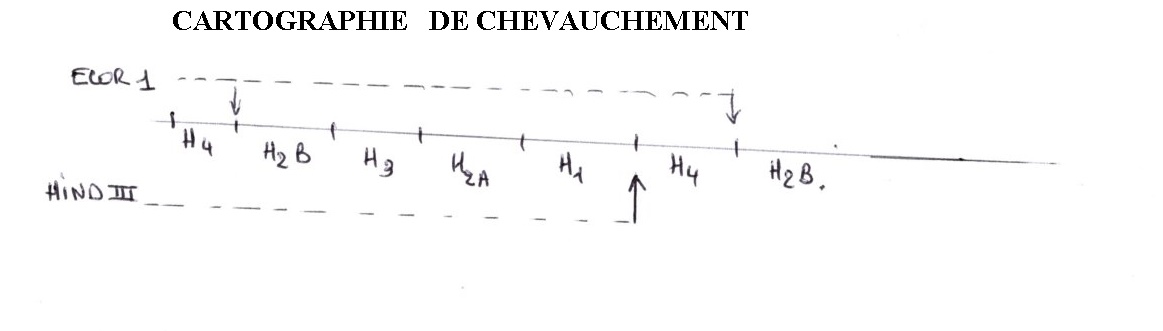
Des enzymes de restriction vont couper entre les gènes:
-ECOR1 coupe toujours au niveau d'une séquence cible entre H4 et H2B. On obtient un fragment avec 5 Gênes.
-HINDIII coupe entre H1 et H4 des fragments qui sont de la même longueur qu'avec ECOR1 et donc dans la même classe.
En faisant agir les deux enzymes, on obtient le segment de restriction H4.
Au moment de l'hybridation, on s'aperçoit que la taille du segment de restriction est supérieure à celle de l'ARNm. il n'en pas de même pour H1;H2b;H2a;H3.
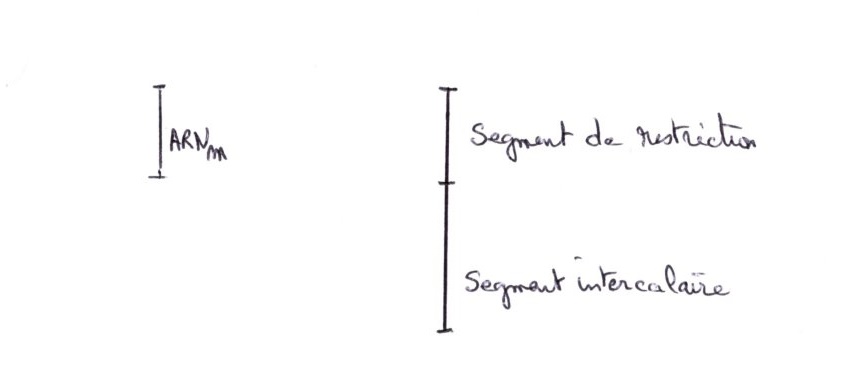
Les gènes Histones comportent donc des séquences non transcrites dans les ARNm matures. Ce sont des segments intercalaires riches en base A & T, qui sont absents de l’ARNm, alors que des portions transcritent sont riches en base G et C.
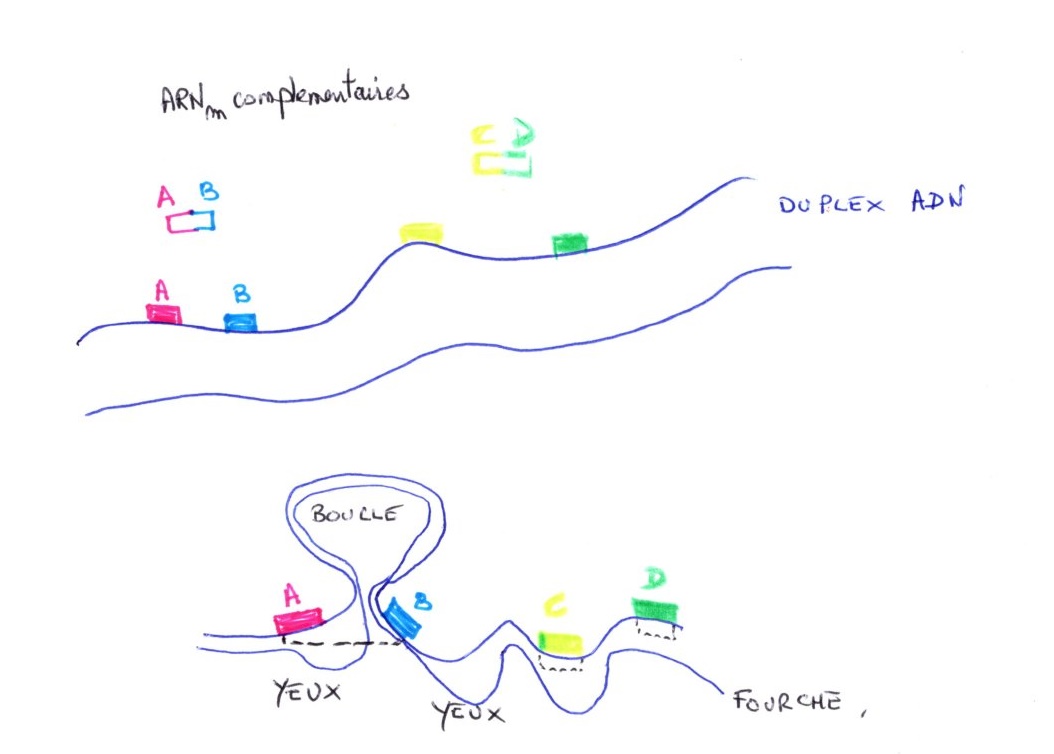
On peut effectuer une dénaturation partielle et une observation au microscope électronique.
Dénaturation partielle
Elle consiste à chauffer modérément ce qui permet de ne pas séparer les bases a été. Pour empêcher la formation des liaisons hydrogène, on utilise la formadine qui se fixe sur A. Il y a ensuite renaturation des régions ne contenant pas en ont pas de A & T, car certaines bases G & C se sont séparées dans la manipulation.
Il y a une formation d’yeux correspondants aux régions qui ne sont pas renaturées, le motif est régulier : œil, segment droit, œil,...
C/ Les gènes uniques :
Le gène étudié est le gène ovalbumine. Ces gènes uniques sont les plus nombreux, ils codent les protéines marqueurs de la différenciation cellulaire. L'ovalbumine est synthétisée par les cellules de l'oviducte de la poule. La quantité de molécule d'ovalbumine est très importante et n'existe qu'un seul gène. Ce gène est très actif car le messager de l'ovalbumine représente 50 % des messagers produits.

Observation microscope électronique :
Organisation en mosaïque du gène:
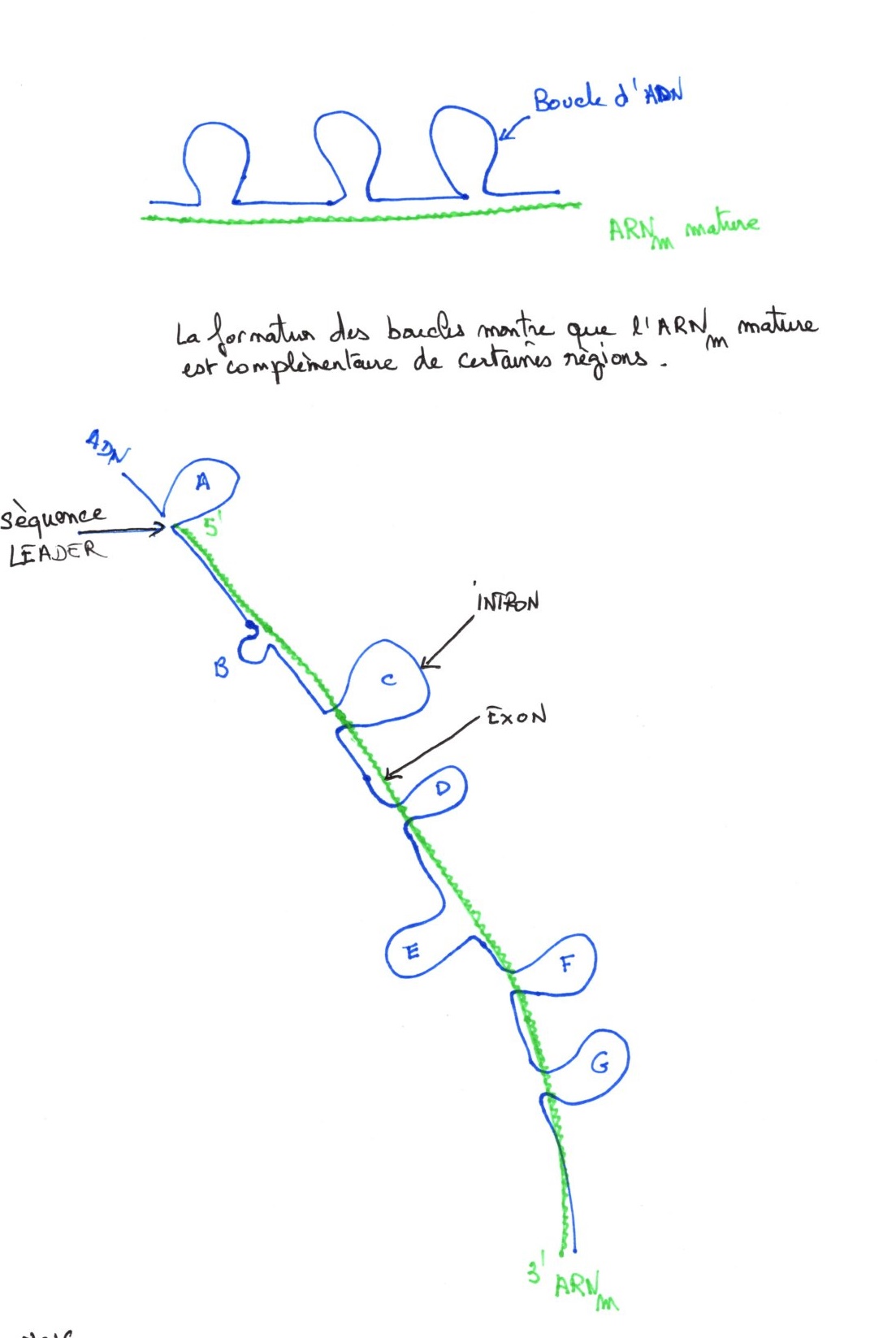
L'ADN (segment de restriction) forme après hybridation, une très grande boucle A, puis des boucles de tailles différentes. L'ARNm s'hybride, en partant de la grande boucle, par son extrémité 5'. La séquence "leader", L, contient les signaux de début de l'ARNm.
Les lettres correspondant aux introns, les zones complémentaires et hybridées à l'ARN messager (chiffres), sont des exons.
C'est la structure en mosaïque car le gène est une mosaïque d'introns et d'exons, de tailles différentes et seuls les exons se retrouvent dans l'ARNm. Intermédiaire entre l'ADN et l'ARN messager, l'ARNm précurseur comprend les introns et les exons. Les introns sont ensuite excisées pour donner l'ARNm mature, qui passe dans le cytoplasme. Après excision enzymatique, il y a épissage des deux extrémités. Les introns sont libérés et dégradés tout de suite dans le noyau. Le pré-messagers est le seul ARN de même taille que l'ADN.
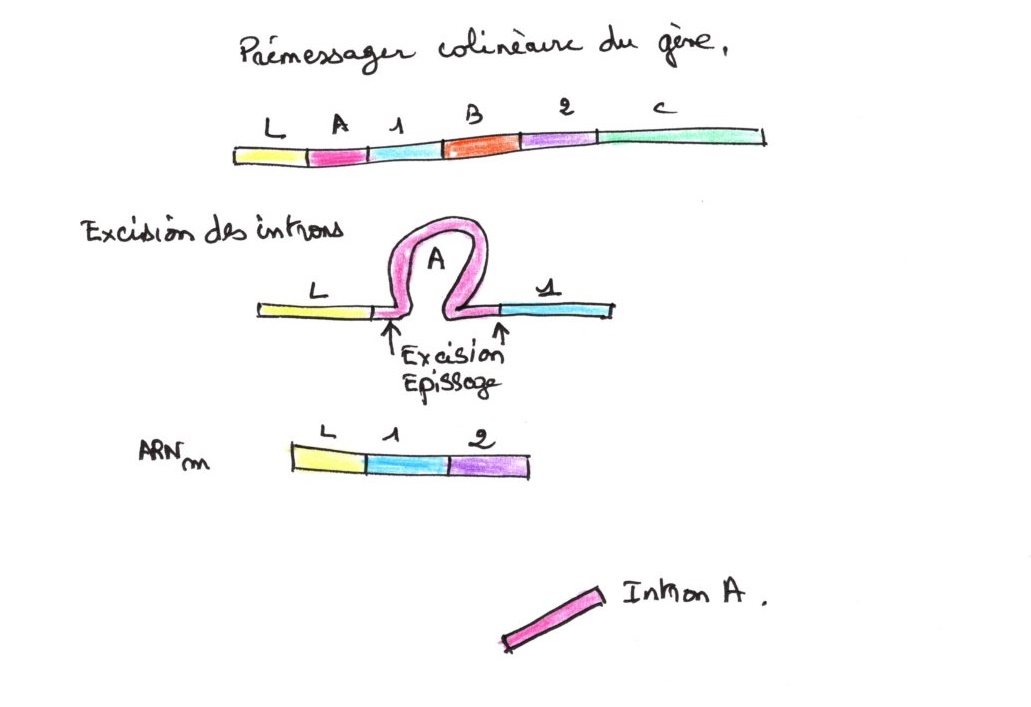
6/Représentation des deux types de chromatines:
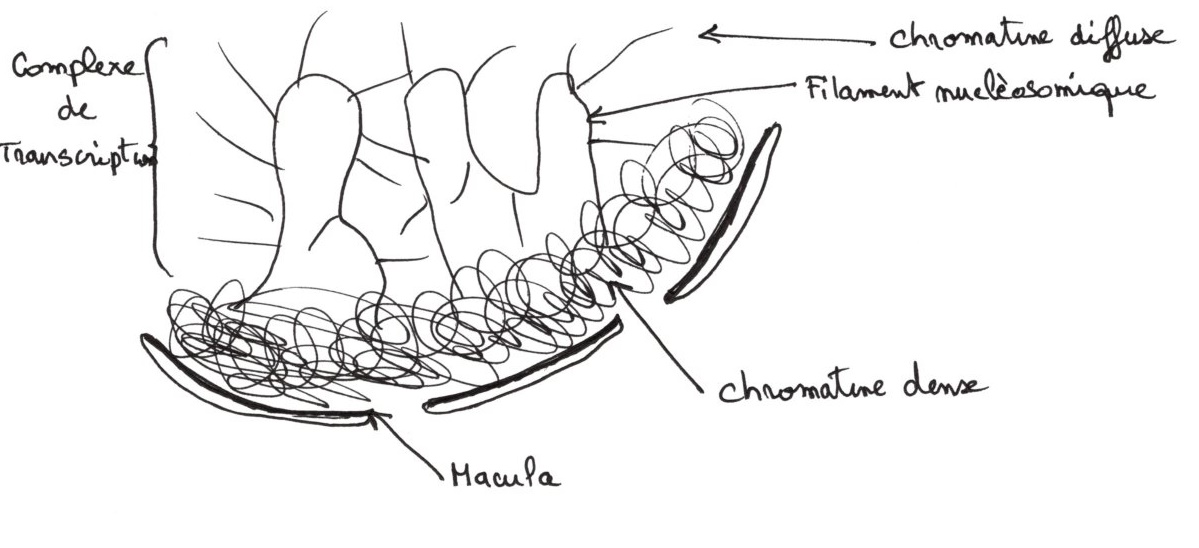
La chromatine peut-être plus ou moins repliée suivant les régions :
- hétérochromatine : est une chromatine dense avec des régions silencieuses pour les transcriptions.
- euchromatine: est une chromatine diffuse avec le transcription de l’ARNm dans les complexes de transcription.
Il y a accumulation de protéines non histones dans l'hétérochromatine.
7/Formation des chromatides :
Les chromatides résultent du dédoublement du matériel nucléaire. en phase S, on peut observer par contraste négatif ou ombrage, les yeux de réplication et il s'agit du dédoublement du filament nucléosomique à certains endroits.
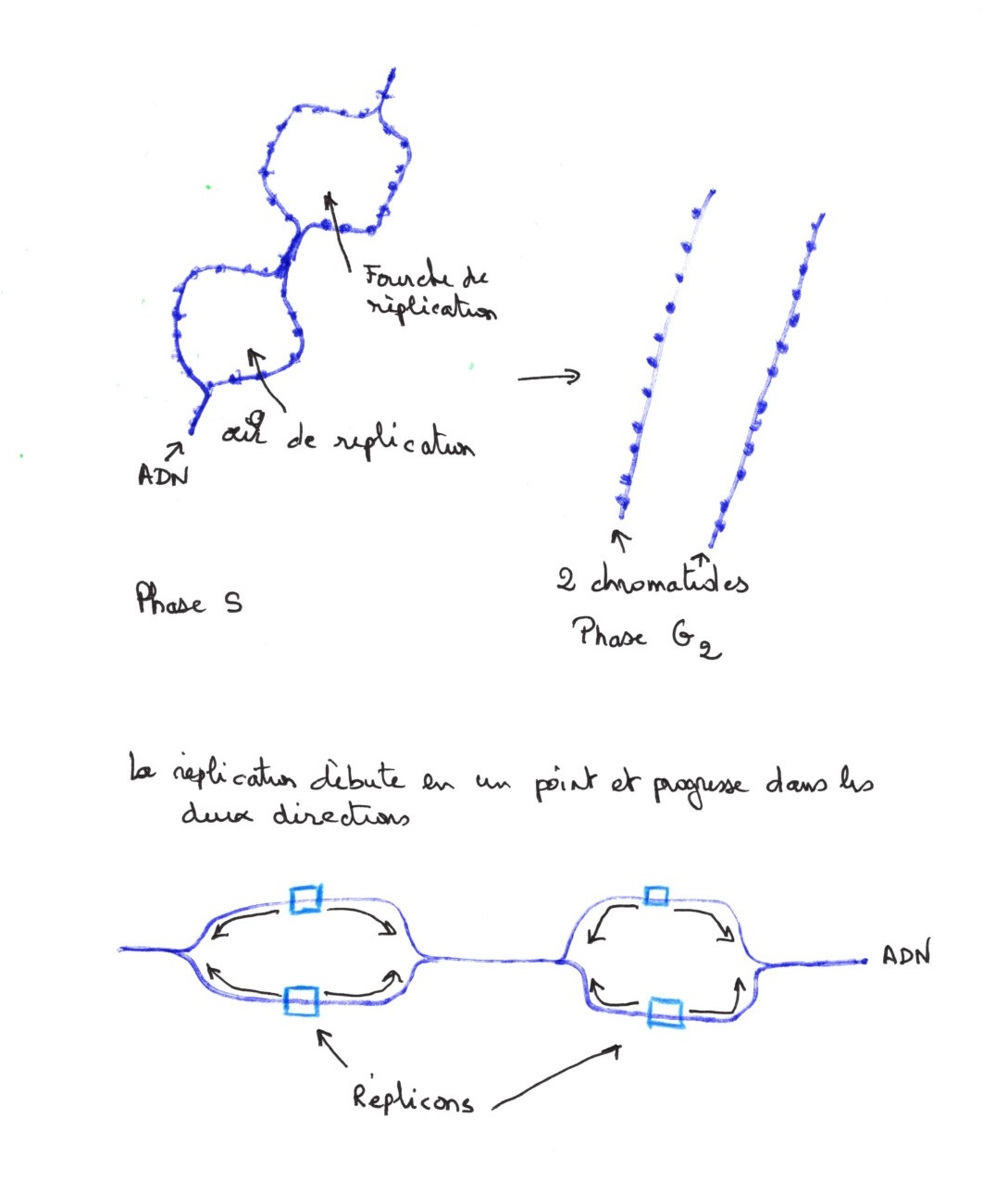
Le réplicon représente le site d'initiation de la réplication de l'ADN. Il y a plusieurs sites d'initiation sur un brin, car il y a plusieurs yeux. Le niveau de la fourche représente le lieu instantané de la réplication au moment de l'observation. Il progresse jusqu'à rejoindre une autre fourche. La vitesse de progression et d'environ un micromètre par minute.
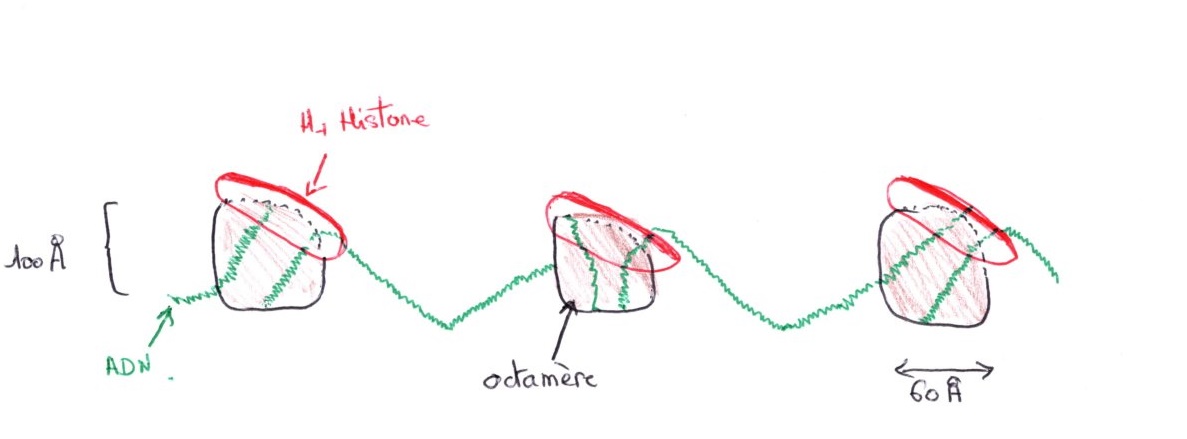
Formation de nouveau nucléosome :
Dès que les nouvelles molécules d'ADN sont synthétisées, il y a fixation des histones et H1 se fixe en dernier pour verrouiller l'ADN. On ne trouve jamais d'ADN dénudé. Dans les embryons en segmentation, il y a beaucoup plus de réplicons que dans les cellules adultes mais ils sont plus petits. La réplication est sous le contrôle de facteurs cytoplasmiques. Pendant la phase G2, la chromatine est dédoublée, cette phase est courte pour les cellules somatiques.
1/ Technique d'analyse de la cellule
5/ Appareil de Golgi ou Dictyosome
13/ Complément sur la méthodologie de recherche en biologie cellulaire
![]()