La membrane plasmique:
C'est l'interface entre l'intérieur et l'extérieur de la cellule ; elle règle
tous les échanges entre le milieu intracellulaire et le milieu extracellulaire;
elle n'est pas visible au microscope optique, mais au microscope électronique.
1/ Composition chimique :
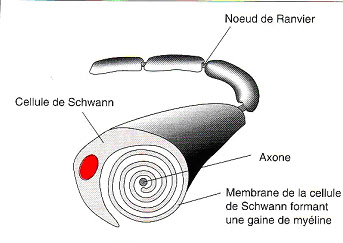
Il faut une grande quantité de membrane pour obtenir une analyse correcte. Le sang contient des hématies(ou globules rouges) qui ne comportent ni noyau, ni organite cellulaire et également la gaine de myéline (Cellule de cellules de Schwann qui entoure les axones des neurones ) sont deux sources de membranes importantes mais, elles sont de structure différente et les conclusions de leur étude ne sont pas toujours généralisables.
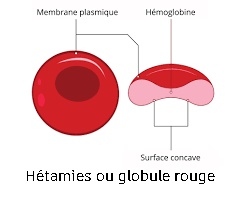
Exemple : les globules rouges:
On les lave de toute trace de sérum ou de plasma, dans une solution isotonique de 9 g de NaCl (0,15 moles), correspondant à 0,3 osmoles de pression osmotique.
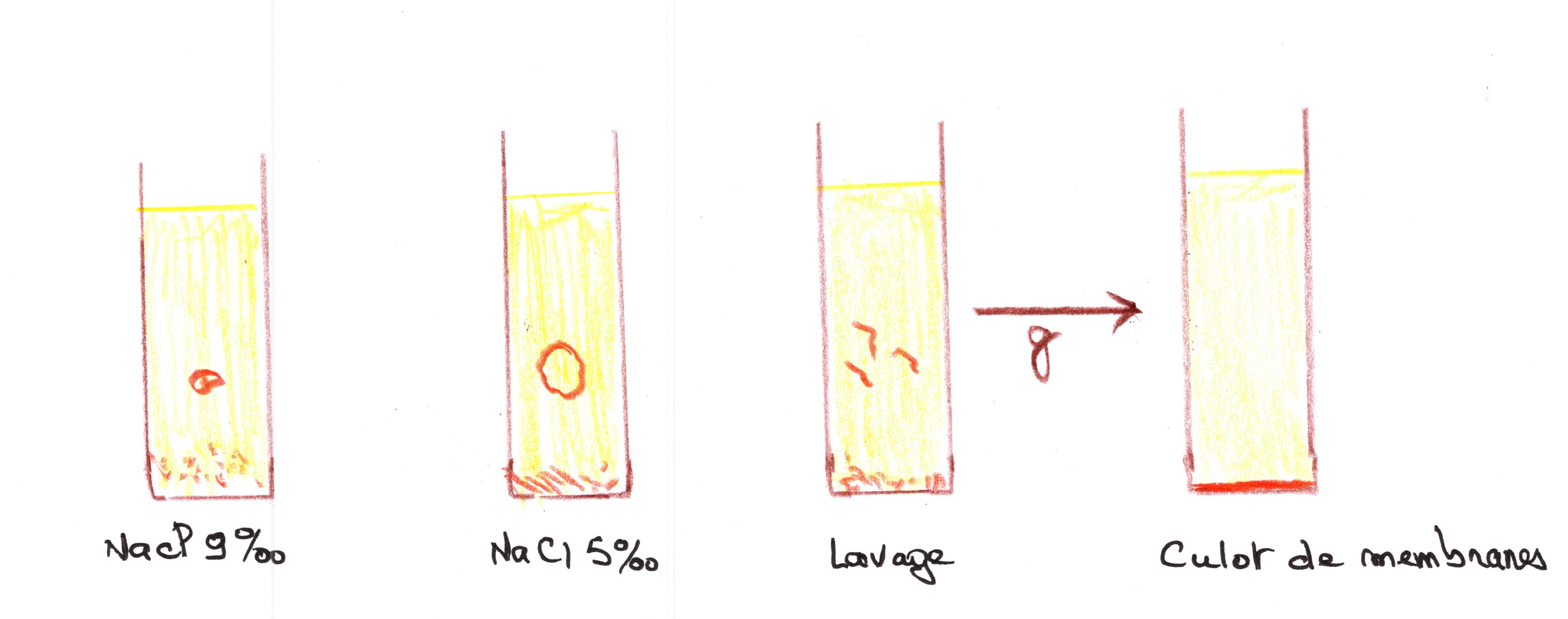
Les hématites sont ensuite éclatées pour séparer la membrane de la cellule par un choc isotonique à 5 g de NaCl. En effet, les cellules se mettent à gonfler puis éclate sous la pression osmotique. L'hémoglobine passe en solution avec le cytoplasme : il y a “laquage” du sang, Il reste alors la membrane plasmique qui est séparée du cytoplasme par centrifugation à 100000 g environ pendant une heure. Le culot de membrane est "resuspendu" dans une solution de NaCl puis centrifugé pour être lavé. Après plusieurs litres de sang, on obtient plusieurs grammes de membrane dont la composition est :
- 50 à 60 % de protéines
- 35 à 45 % de lipides
- 1 à 3 % de glucides
A/ Lipides:
Ils sont isolés dans l'alcool éther qu'il solubilise, on peut ainsi étudier les différents lipides : phosphorés et non phosphorés.
Les phospholipides :
Les phosphoglycériques:
Il contient du glycérol et à la double propriété d'être soluble dans les corps gras et dans l'eau.
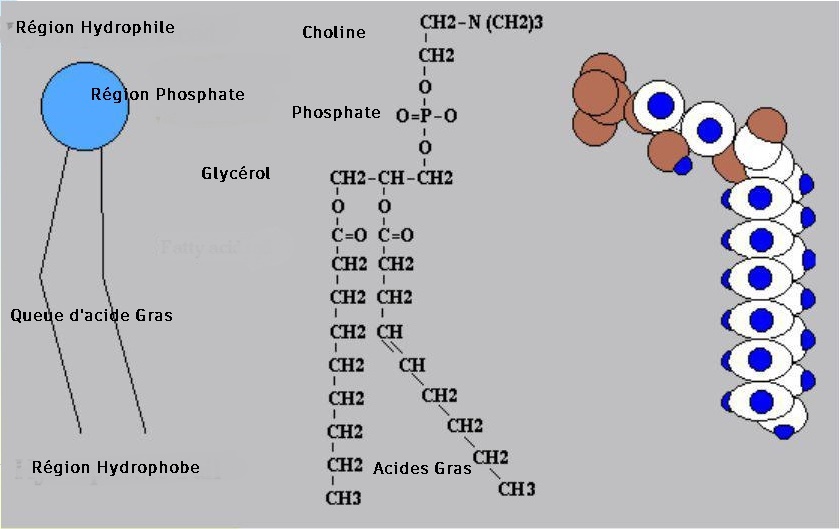
Exemple de phosphoglycérides : La lécithine est composée d'acide phosphorique, d'un amino-alcool : La Choline, d'un glycérol, et de deux acides gras : L'acide palmitique et l'acide oléique.
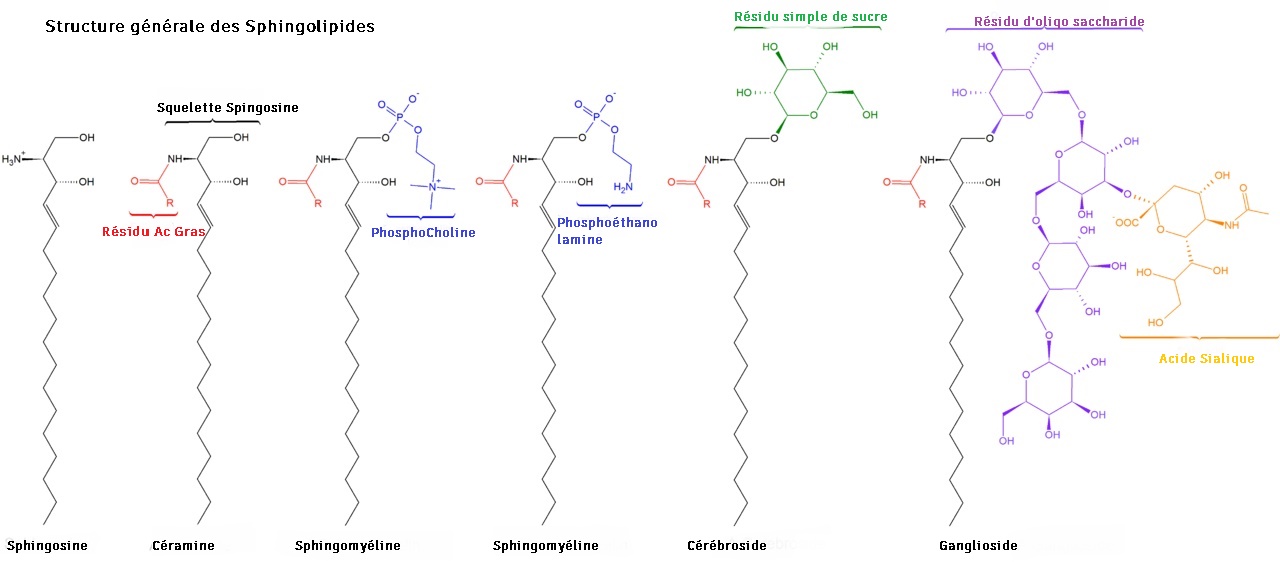
Les sphingolipides :
On les trouve dans les tissus nerveux où le glycérol est remplacé par la sphingosine
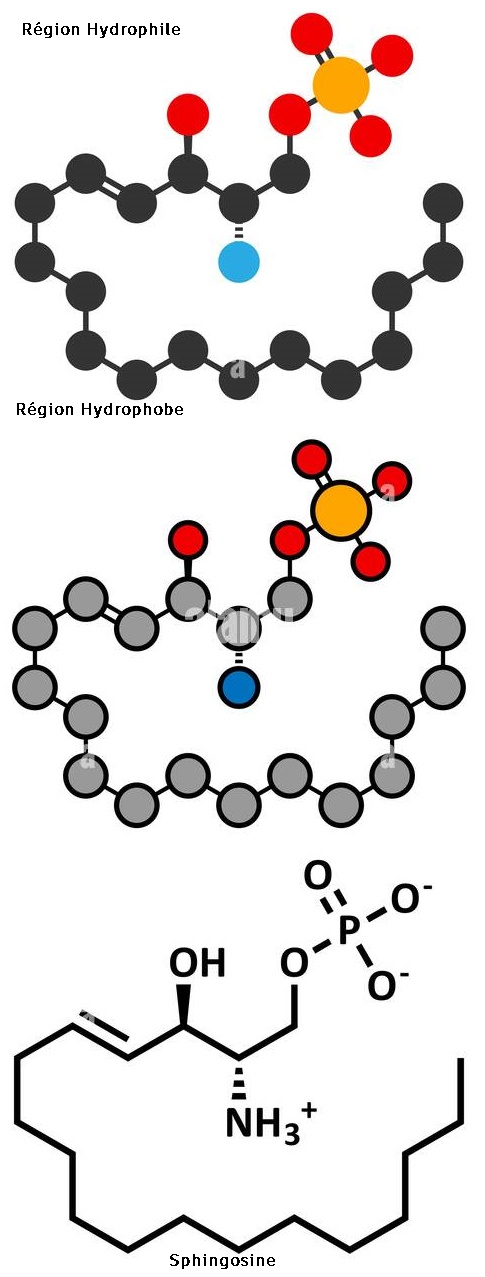
Si l'amino-alcool est la Choline, il s'agit de la sphingomyéline.
Les Lipides non phosphoryles :
Les glycolipides :
Ce sont des lipides liés à des glucides, présents en grande quantité dans les gaines de myéline. Il comporte:
Sphingosine + Acide Gras + Glucide
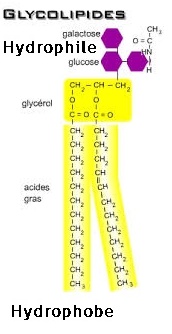
À noter que si la tête glucidique est un hexose, il s'agit alors d'un cérébroside et si la tête est un oligo-saccharide, il s'agit d'un ganglioside.
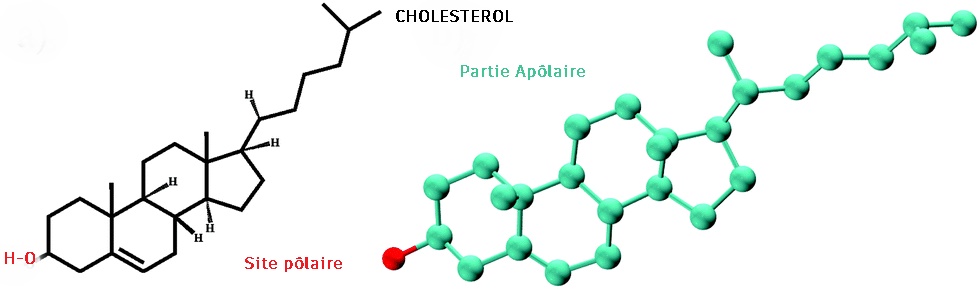
le cholestérol :
Il n'est présent que dans les cellules animales. Le cholestérol membranaire augmente parallèlement avec l'augmentation de l'eau dans les membranes cellulaire..
B/Les protéines :
On en distingue deux types en fonction de leur propriété :
les protéines de structure ; à l'état figuré, elles sont observables.
les protéines enzymatiques. Malgré tout, cette distinction est plutôt arbitraire. Par exemple: La myosine est structurale et à la fois enzymatique car ATPasique (intervient dans la contraction musculaire). On peut d’ailleurs la séparer. Cette protéine permet de libérer de l'énergie en hydrolysant l'ATP en ADP.
Les glycoprotéines sont des protéines de surface, souvent antigéniques. exemple ci-dessous : groupe sanguin ABO.
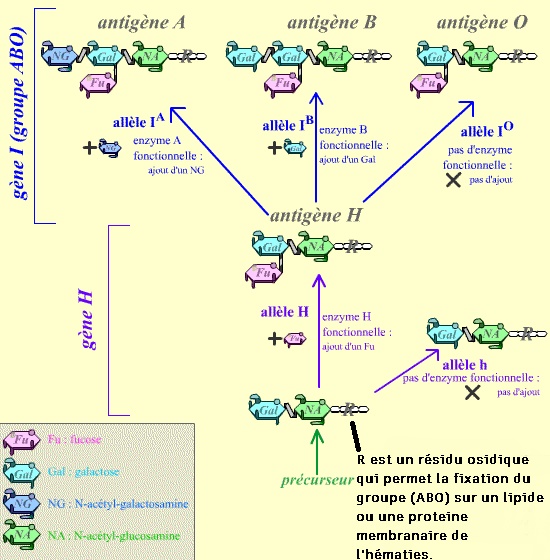
Les allèles d'un gène permettent la grande diversité génétique entre les différents individus. Les allèles peuvent coder un caractère physique ici pour le groupe sanguin, mais par exemple, la couleur des cheveux, des yeux,...
2/ Organisation moléculaire:
Il s'agit d'un modèle physico-chimie, dont le point de départ et la composition chimique de la membrane avec les phospholipides arrangés entre eux.
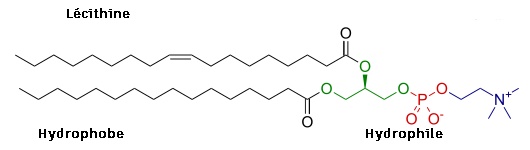
Ac.Oléique + Ac.Palmitique + Glycérol + Phosphate + Choline
Globalement, pour la membrane plasmique, on sait qu'elle sépare deux milieux très hydratés. Les parties hydrophiles sont orientées vers l'extérieur et l'intérieur de la cellule. Il y a donc une bicouche phospholipidique.
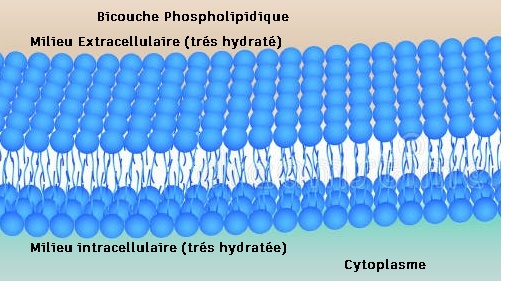
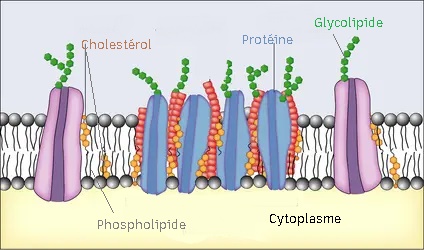
La membrane a une certaine rigidité : la cohésion des molécules est due à des interactions de type électrostatique entre les molécules de même nature, les interactions hydrophobes sont tournées vers l'intérieur de la membrane, quant aux protéines , elles se placent au niveau des pôles hydrophiles c'est-à-dire vers l'extérieur de la membrane.
Modèle de Danielli et DAWSON (1935):

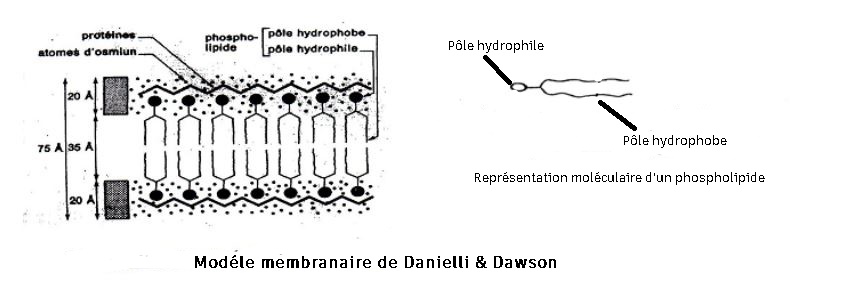
Mais avec la microscopie électronique les observations de Robertson en 1952 permettent de voir suivant la technique de fixation les structures rendues denses aux électrons par l'oxyde d'osmium (OsO4) qui insolubilise les protéines.
Photo d'un microscope électronique et schéma de principe:
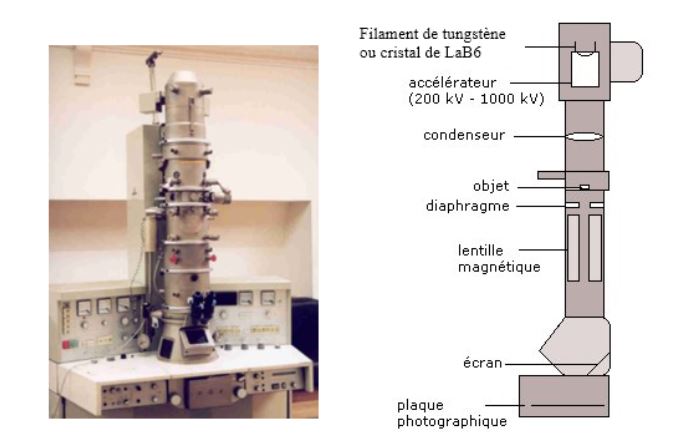
On peut voir trois couches distinctes, déjà prévu par Robertson.
Sur ce schéma le contraste est dû à la présence des métaux lourds dense aux électrons, le tétraoxyde d'osmium crée un contraste en se fixant sur les protéines et les phospholipides.
Lors du protocole,les protéines réduisent l'osmium, elle représente deux couches externes de la membrane. Robertson a montré que toutes les membranes cellulaires ont la même structure tri lamellaire d'où la notion de membrane unitaire. Cette recherche a été complétée par Singer et NIcholson et a abouti au schéma ci-dessous.
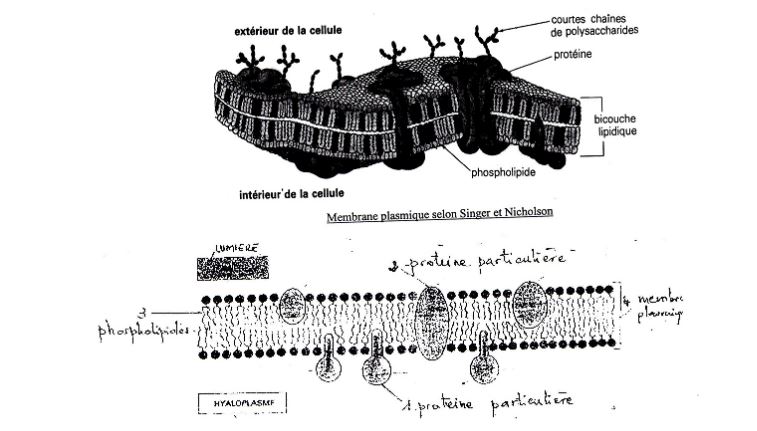
a) Confrontation du modèle et de l'observation avec les propriétés de la membrane plasmique:
Les modèles observés n'arrivent pas à expliquer la perméabilité de la membrane cellulaire aux substances aqueuses et produits gras qui devraient être arrêtées par les parties hydrophobes.
On retrouve cette même structure organisationnelle pour toutes les membranes quelles soient mitochondriales, de Golgi, réticulienne endoplasmique,... mais elles ne sont pas perméables aux mêmes substances.
Il faut donc pour le comprendre effectuer de nouvelles observations en créant de nouvelles techniques.
- la double fixation:
Pour mieux un insolubiliser les constituants de la membrane, on effectue un tannage par la glutaraldéhyde. Après un rinçage par l'OsO4, ce dernier est réduit au niveau des protéines déjà insolubilisées. L’observation permet de voir une discontinuité de la membrane.

La membrane apparaît comme une mosaïque faite de sous unités. On peut noter une différence entre les autres sous unités de la membrane plasmique et d'autres membranes intracellulaires tel que le réticulum endoplasmique, la membrane des mitochondries, le Golgi,...
- Le Cryodécapage:
Cette méthode permet d'observer la surface membranaire. mais, comment disséquer une membrane ?
En résonant en toute logique sur l'existence certaine du squelette phospholipidique : la partie la plus solide étant celle qui a le plus d'interactions électrostatique, donc la cohésion plus grande. On peut en déduire que la partie la moins solide se situe au niveau des zones hydrophiles. Quand on congèle à moins 190 degrés, il y a inversion des zones de rigidité de la membrane, les zones hydrophiles sont beaucoup plus solides, les zones hydrophobes ne changent pas. Dans ces conditions, il y a un plan de clivage naturel de la membrane entre les couches de phospholipides.
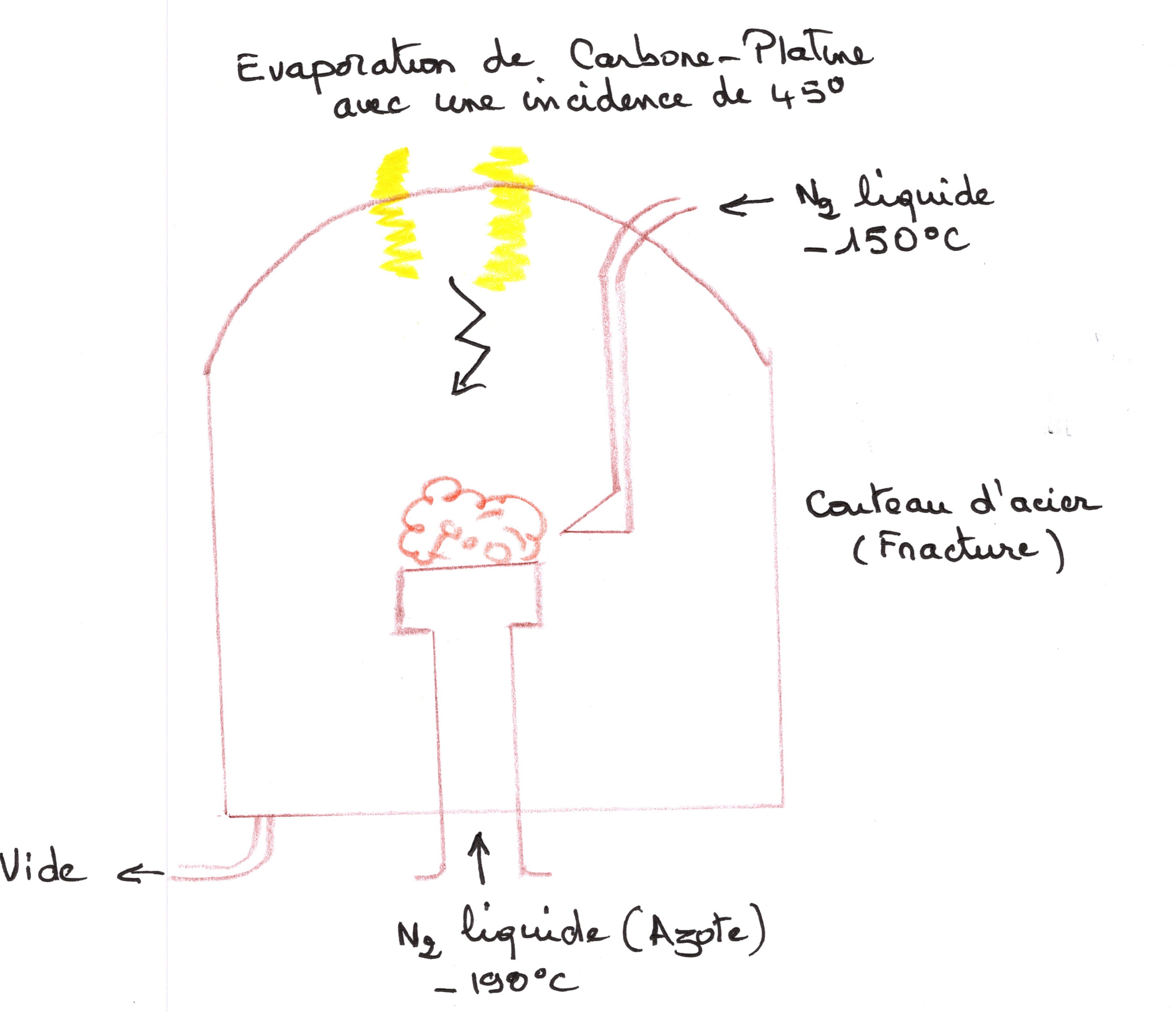
En avant du tranchant du couteau, il se produit une micro fracture. Le cytoplasme est peu fracturé, il est rendu dur, du fait qu'il est très hydraté, la fracture passe donc entre les deux couches de phospholipides. Un arc électrique vaporise la matière des électrodes : carbone et platine. Il y a un nuage d'atome produit sous vide. La surface du tissu reçoit des vapeurs de carbone et de platine.
Le dépôt de carbone platine ne se fait que sur des endroits exposés à la vapeur. Il s'agit d'une “réplique transfert” qui contient l'information par un phénomène d'ombrage. Ensuite, tout le tissu est dissous sauf la réplique transfert qui en représente le moulage.

Si il n'y a pas de protéine, la surface observée est complètement plane, car les images de creux et de relief représentent les emplacements de protéines.
La distinction entre creux et relief se fait grâce au principe d'inégal épaisseur du dépôt de vapeur, et par la connaissance de la direction de l'ombrage.
b/ Structure en mosaïque : le Cryodécapage et la double fixation permettent d'arriver au modèle actuel de membrane. Cette structure tient compte de la fluidité de la membrane (mobilité dans un même plan).
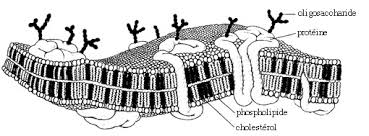
Les protéines ont des propriétés différentes. Une protéine partiellement intégrée possède des caractéristiques hydrophobes. toute cette structure est fluide et mobile renouvelée en permanence.
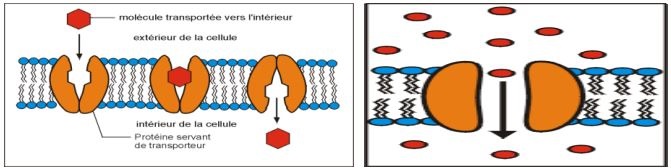
Le squelette de la membrane est une bicouche phospholipidique sur laquelle s'intègre des protéines différentes, expliquant la perméabilité membranaire.
Modèle micellaire : Permet de comprendre comment se forme un pore dans la membrane et capable de fixer des ions. Dans le modèle micellaire, il y a orientation concentrique par fluidification, avec équilibre par pression réciproque. Ce motif est hexagonal et ainsi il y a création d'un pore.
c/ caractère du modèle en mosaïque : la symétrie.
On se propose de démontrer que les protéines ne sont pas les mêmes à l'extérieur qu'à l'intérieur de la cellule. le principe de cette expérience repose sur le marquage par la radioactivité des protéines membranaires. les protéines extérieures sont marquées par iodination avec de l'iode 135 .
Il ne faut pas que le marqueur traverse la membrane, l'iode 135 est fixé par lactoperoxydase sur les résidus de tyrosine. la lactoperoxydase reste extracellulaire, elle ne traverse pas la membrane. le globules rouges est une source commode de membrane plasmique, on utilise des fantômes.
les vésicules obtenues sont iodinées, elles sont mises en présence diode 135 et de lactoperoxydase (protéines enzymatique de haut. moléculaire que notre traverse pas la membrane) .
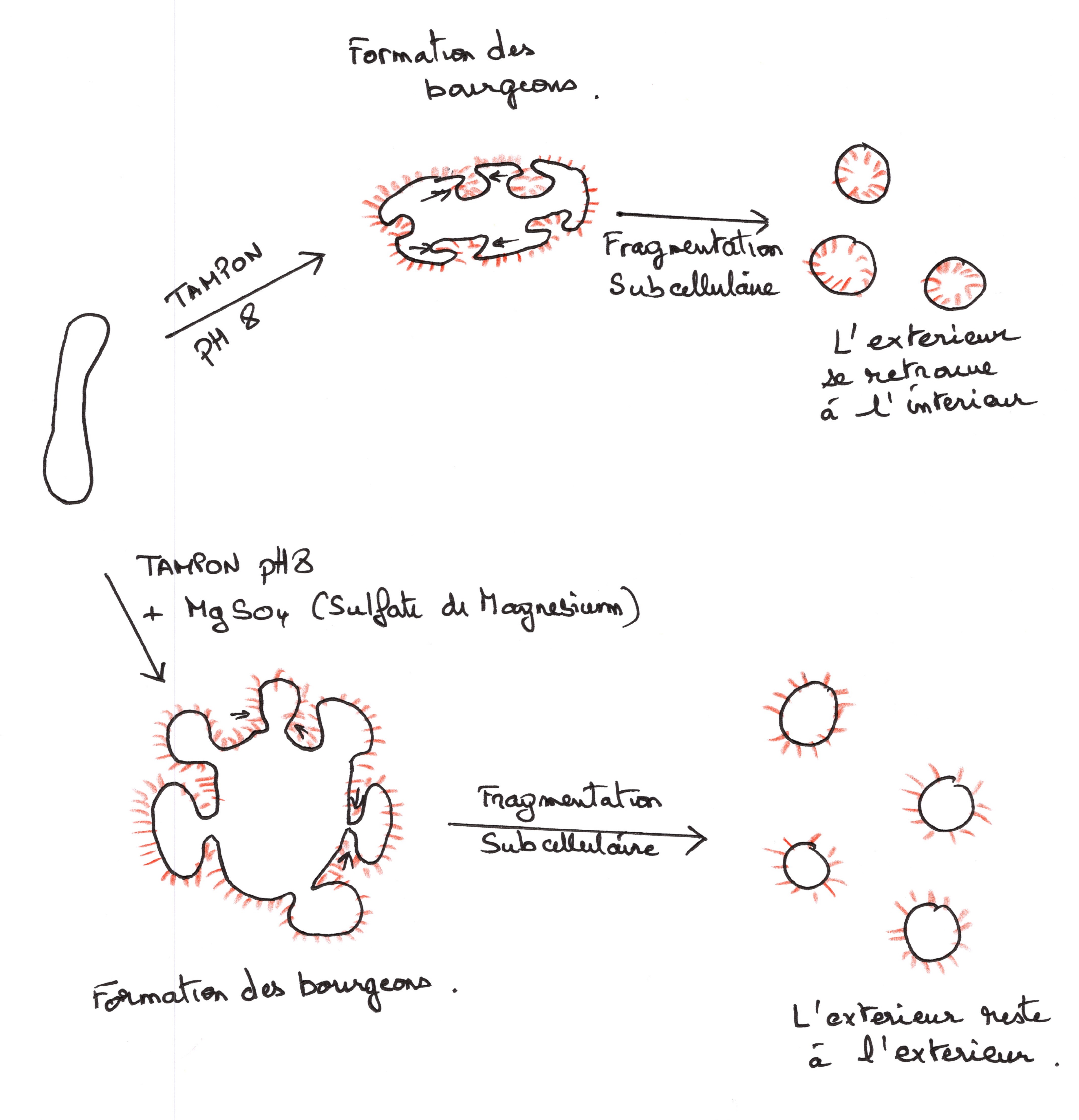
Seul le côté externe des vésicules sera marqué par l'iode radioactif. On sépare
ensuite les protéines des autres constituants membranaires. Les protéines sont
séparées par électrophorèse, puis on verse de l'émulsion photographique (Nitrate
d’Ag) pour révéler la radioactivité. En comparant les électrophorèse des deux
types de vésicules, on peut voir que les bandes de protéines sont identiques
mais que la radioactivité y est inégalement répartie.
Nous pouvons conclure de cette expérience que ce ne sont pas les mêmes protéines
qui sont marquées, suivant que nous considérons les protéines sur les sacs de
l'intérieur ou de l'extérieur ; mais un troisième type est révélé, elles sont
présentes dans les deux types de vésicules, elle traverse la membrane et
effleurent le côté externe et le côté interne.
S’'il y a des protéines qui traversent la membrane, elles doivent comporter une
partie hydrophobe au niveau des phospholipides.
Exemple dans les globules rouges où la glycophorine a pu être isolée, purifiée
puis séquencée.
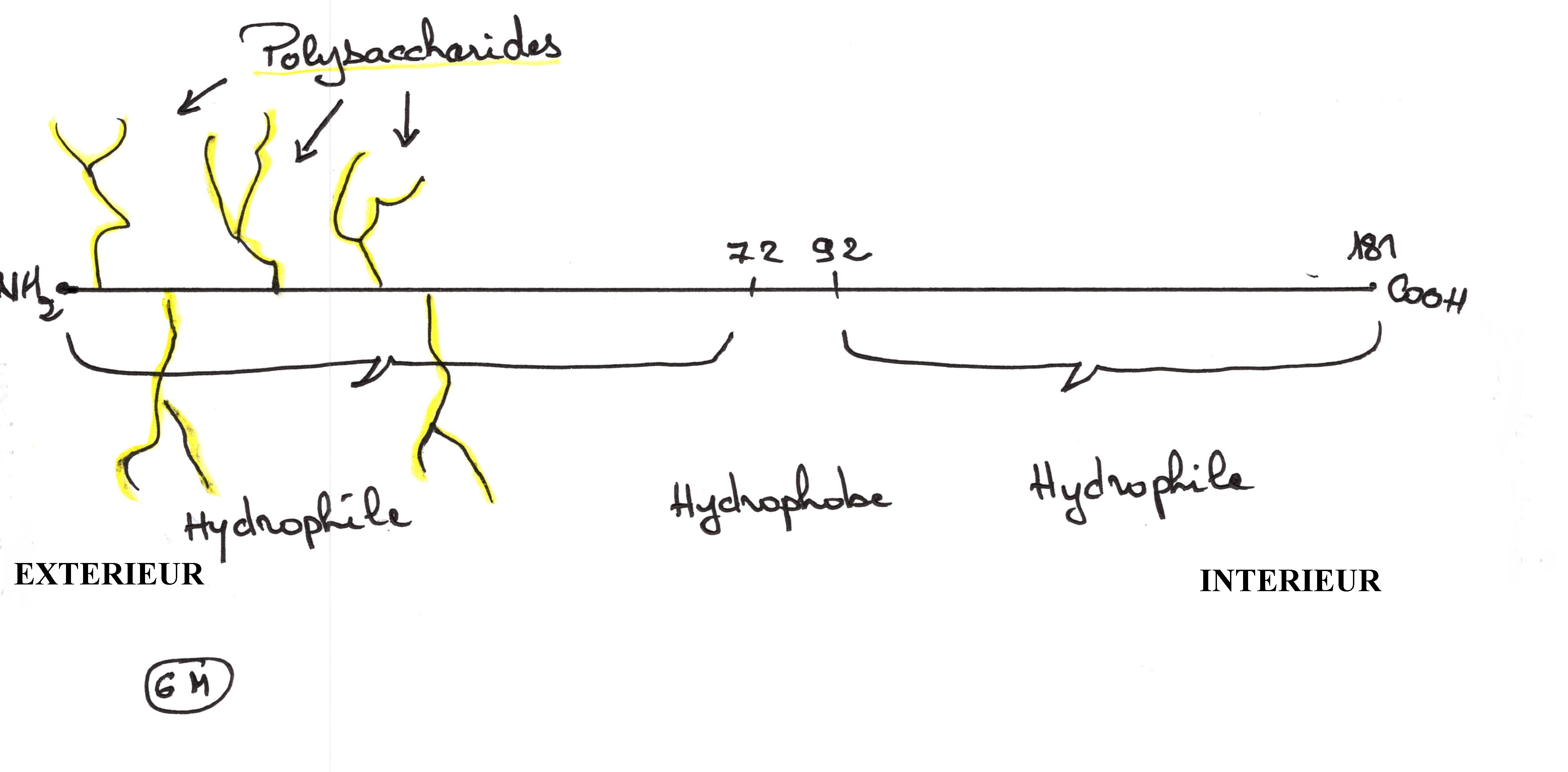
Après analyse, on a pu trouver 16 résidus polysaccharidiques qui sont fixés dans
la première partie, vers l'extérieur, ils sont constitués d'acide sialique et
d'oligosaccharide.
Sur la chaîne protéique de 72 à 92 se trouve des acides aminés hydrophobes comme
la proline donnant ainsi des interactions hydrophobes avec les phospholipides.
d/ Deuxième caractère du modèle en mosaïque : la mobilité des protéines dans le
plan de la membrane.
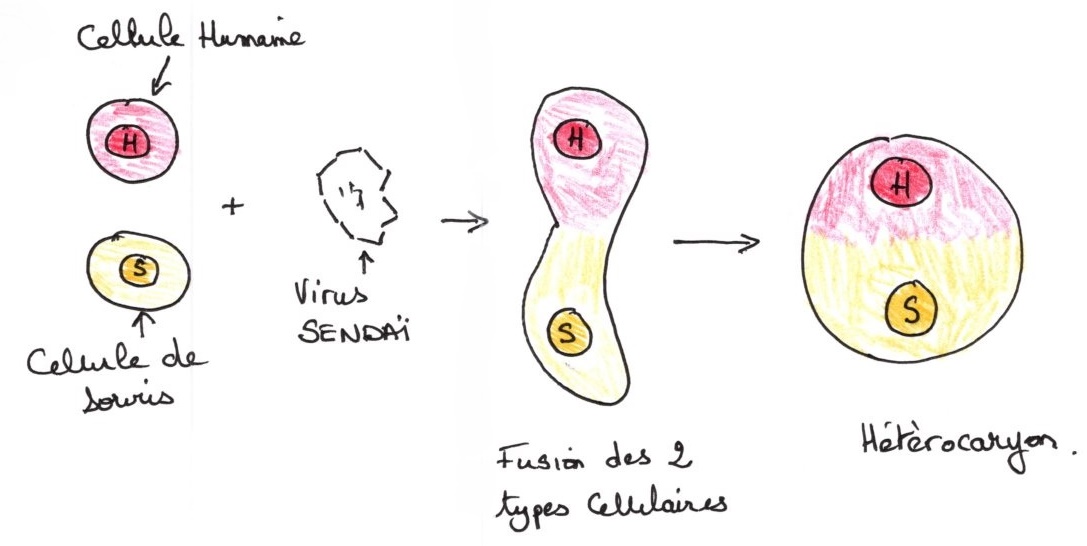
Cette expérience repose sur trois principes : formation d'hétérocarion : ce sont des chimères qui résultent de la fusion de deux cellules d'espèces différentes
Production d'anticorps contre l'antigène de surface porté par les polysaccharides du côté externe de la membrane.
Les anticorps ne sont pas visibles directement, il faut un marquage fluorescent.
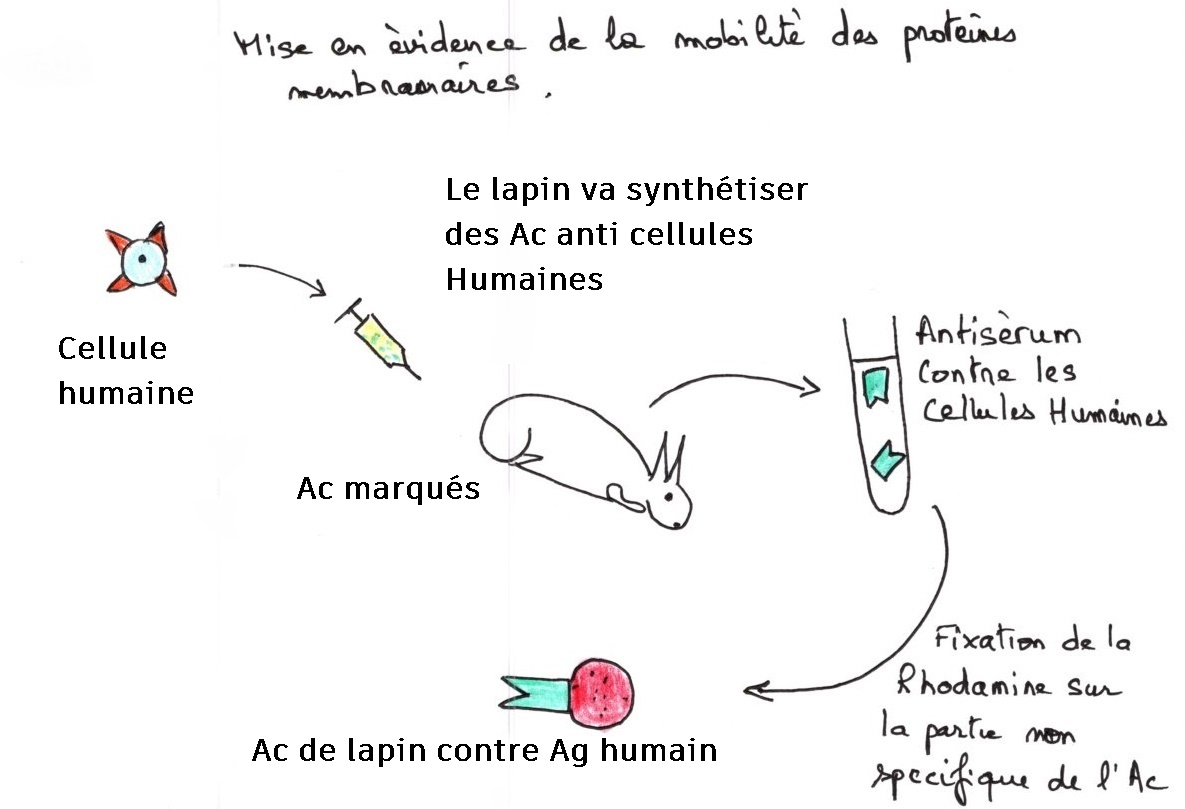
Deuxième étape:
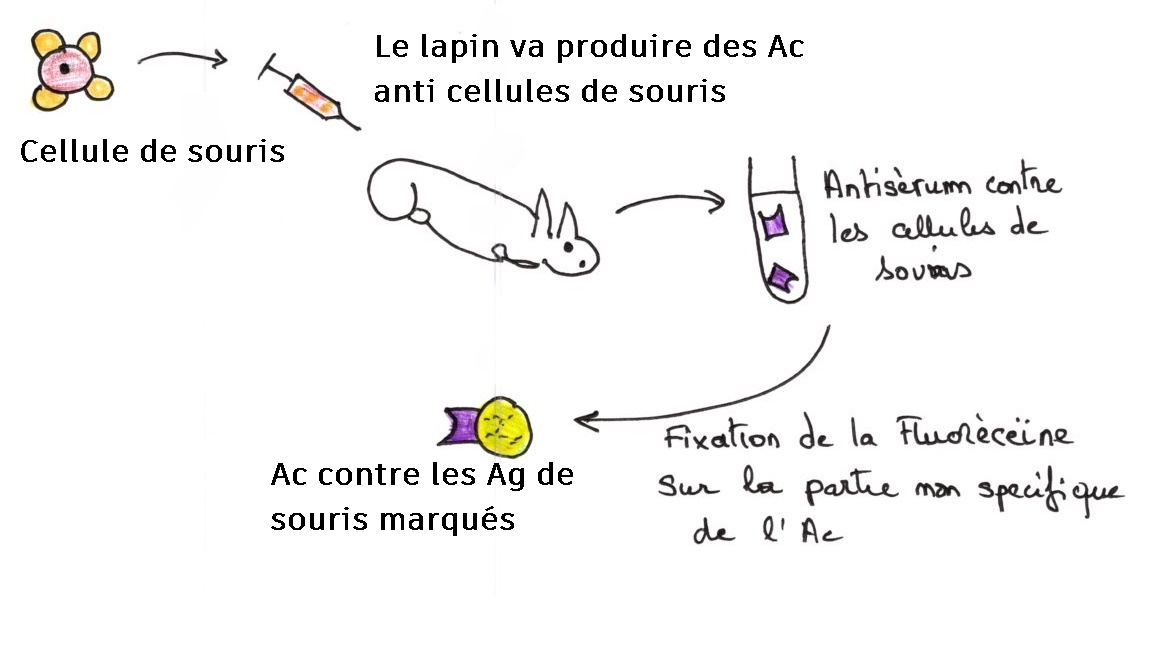
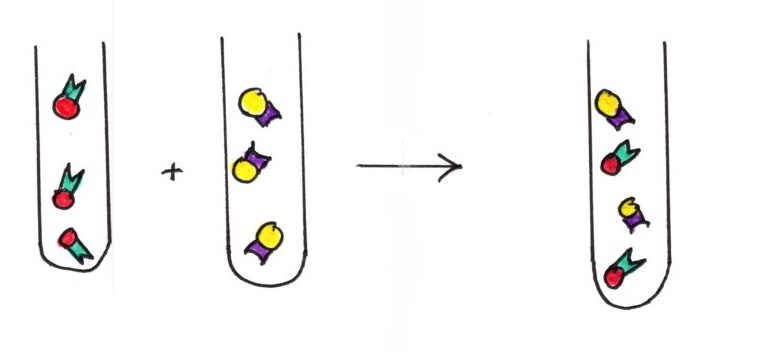
Troisième étape : réalisation de l’hétérocaryon. On mélange les 2 types d'anticorps, marqués différemment.
La présence du virus Sendai provoque la formation de l'hétérocaryon par fusion des membranes et mise en commun des cytoplasme.
A propos du virus Sendai:
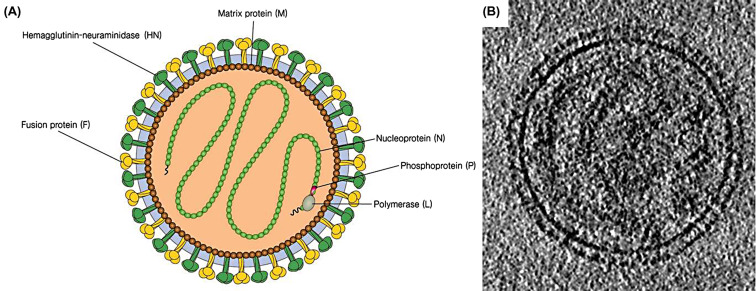
Le virus Sendai est un type de virus à ARN- de la famille des
Paramyxoviridae, découvert pour la première fois chez une souris à Sendai, au
Japon. Il provoque de graves infections respiratoires chez la souris, mais
seulement des maladies bénignes chez l'homme, ce qui le rend adapté à la
recherche. Le virus possède des Ag de surface qui lui permettent de se fixer sur
les cellules humaines et de souris. Il permet la fusion des deux types
cellulaires créant ainsi un hétérocaryon.
Prélèvement toutes les 5 minutes
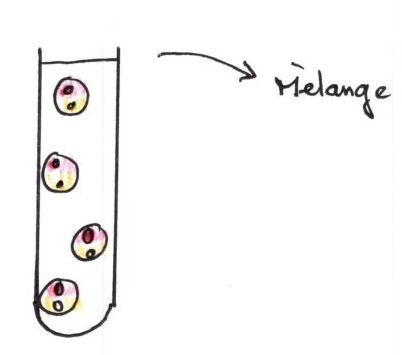
résultats :
5 minutes après contact, par UV, on observe deux territoires membranaires, il y a donc peu de modifications, les territoires sont intactes
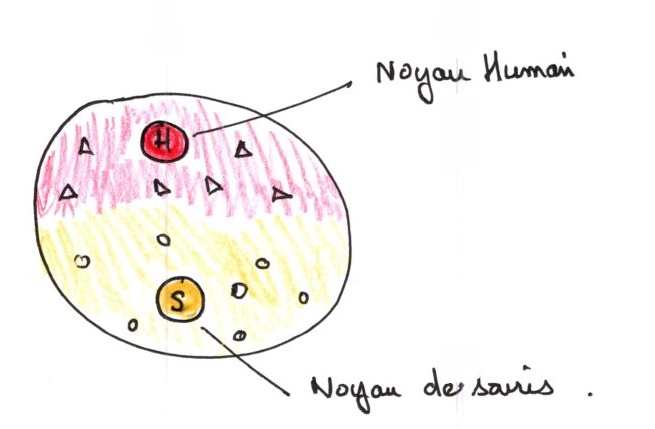
40 minutes après contact, on ne distingue plus de différence de répartition de couleurs en surface membranaire.
Avec un filtre vert, qui arrête la couleur rouge, on voit que la fluorescence verte occupe toute la surface de la membrane, de même pour la fluorescence rouge.

Après 5 minutes, des complexes antigènes anticorps n'ont pas eu le temps de migrer dans la membrane, les territoires sont différenciés.

En 40 minutes, les complexes sont une nouvelle répartition, il y a un mélange complet, donc déplacement des protéines dans le plan de la membrane. Les protéines se déplacent bien en surface de la membrane cellulaire, il y a donc grande mobilité des protéines dans le plan de la membrane.

Autre expérience : comment influencer les déplacements des protéines membranaires?
Exemple : Il s'agit de vérifier la mobilité des protéines dans le plan de la membrane, cette expérience montre une cause de déplacement des protéines.
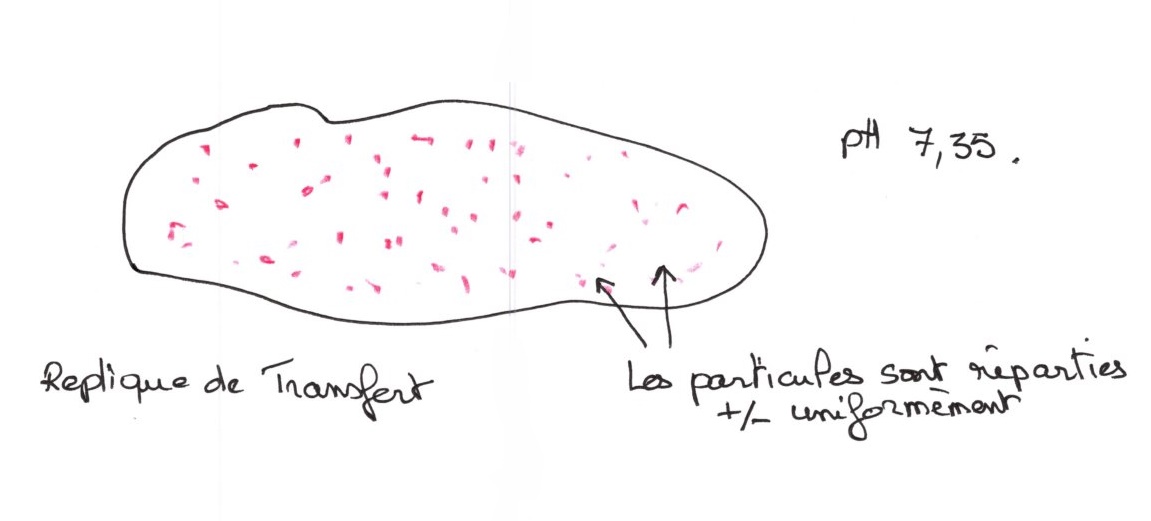
En utilisant la technique de Cryodécapage, des fantômes sont incubés à PH physiologique, puis congelés, cryofracturés. On effectue ensuite l'ombrage et l'observation de la réplique avec l'analyse des particules qui font saillie à la membrane.
Mais en effectuant la même expérience à pH acide, on observe :
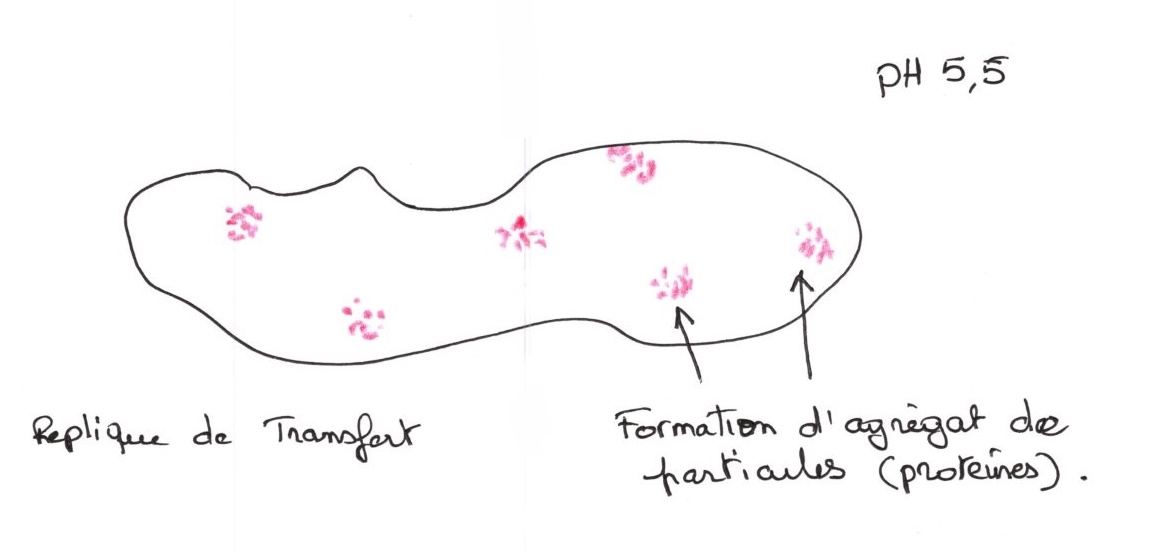
Cette expérience est réversible, si on lave les fantômes entre pH 5,5 et 7,5.
L'implantation des protéines dans la membrane dépend de la répartition des charges au niveau membranaire.
A PH 7,5, l'environnement des charges n'est pas modifié, la mosaïque est régulière avec des protéines intercalées périodiquement.
En augmentant la concentration en h+, il y a agrégation de protéines, elle se rassemble dès que l'on modifie l'environnement des charges.
Ce phénomène est réversible, il donne une conception dynamique de la structure de la membrane.
3/ Le revêtement externe
Le revêtement cellulaire ou cell coat, représente la structure la plus externe de la membrane cellulaire. Elle est constituée essentiellement de polysaccharides et de mucopeptides.
Ci-dessous, l'amibe.
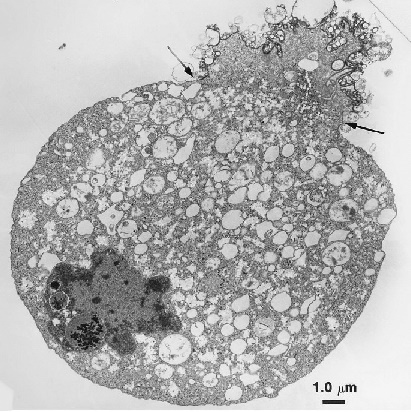
Le microscope électronique permet de mettre en évidence différents types de revêtement cellulaire, par exemple:
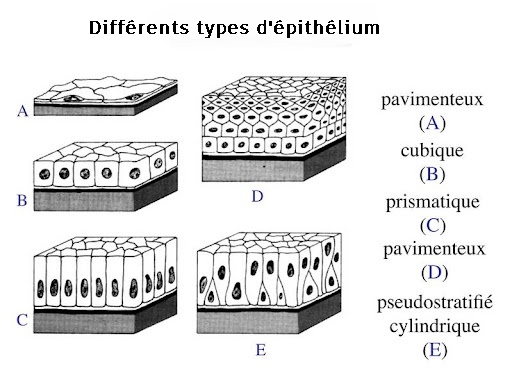
-microvillosité de l'épithélium intestinal et différents épithélium

Revêtement de type cimentaire
-Les desmosomes entres les cellules épithéliales
Revêtement hypertrophié
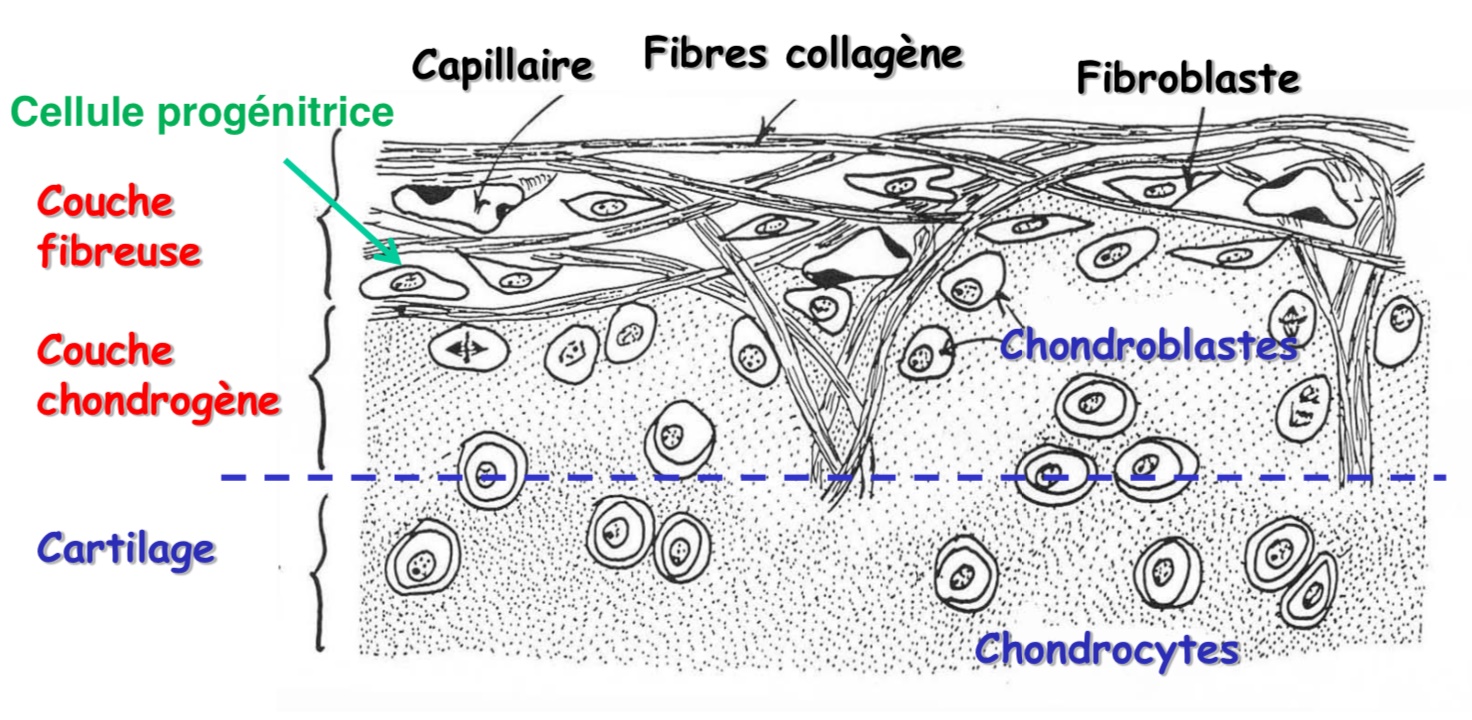
- Le cartilage (Les cellules progénitrice ont la capacité de se différencier en différents types cellulaire, ici en fibroblaste ou chondroblaste car le milieu dans lequel elle se situe et qui contiennent des effecteurs chimiques, physiques,... l'induisent cette orientation cellulaire).
Fusion intercellulaire
- La gaine de myéline (sorte de syncytium)
- Le revêtement fibreux des bactéries (Bacillus) glycocalyx.
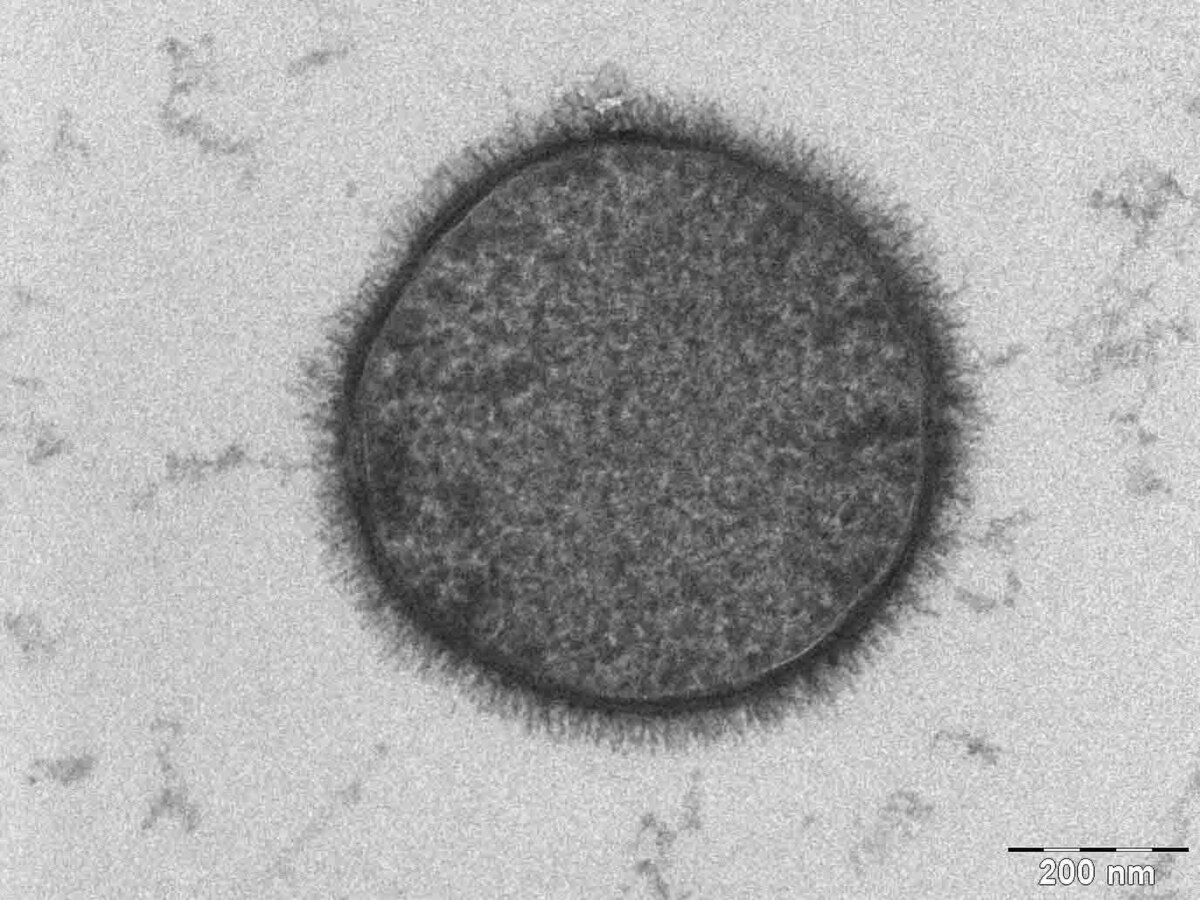
Différents types d'absorptions ou d'excrétions de la cellule.
Le phénomène de pinocytose :
Il s'agit d'une absorption ou d'une sécrétion de gouttes de liquide, sans qu'il y ait contact entre les milieux intracellulaires et extracellulaires.
Exemple : les cellules endothéliales des capillaires sanguins sont le siège de pinocytoses intenses.
Observation statique:
Exemple avec la ferritine et la peroxydase. Après injection intraveineuse, la substance arrive dans la lumière du capillaire puis rentre par les vésicules. On met ainsi en évidence le sens de la pinocytose. Le transport par pinocytose à la caractéristique de ne jamais mettre en contact le milieu intracellulaire avec le milieu extracellulaire.
Mécanisme de l'exocytose :
Il est comparable à celui de l'endocytose qui fonctionne en inverse. Voici le schéma de la membrane plasmique au microscope électronique avec un faible grossissement.
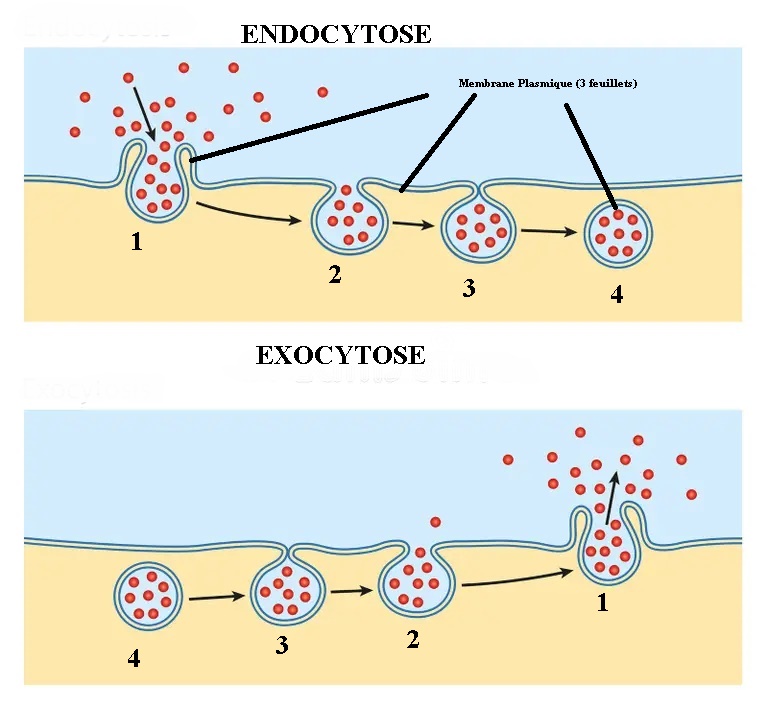
1- Stade d'approche de la membrane
2- stade d'accolement, on observe trois feuillets sombres, il y avoir fusion des feuillets.
3- stade d'ouverture
4- stade d'intégration à la membrane
Constatation:
Il n'y a jamais rupture de la membrane plasmique, qui a une structure
continue. Une fusion précède toute ouverture. La membrane plasmique est
l'interface entre le milieu extérieur et intérieur.
Il
existe aussi d'autres phénomènes plus ou moins passifs d'entrée ou de sorties de
particules tels que les sels minéraux ou petites molécules dans les cellules.Le
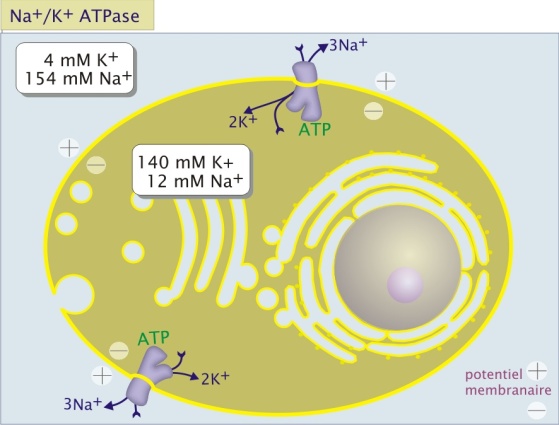
transport peut être actif (nécessite de l'énergie) ou passif. Le plus connu est
celui du Sodium Na+ et du potassium K+.La pompe Na + K + est une protéine
ATlasique transmembranaire électrogène découverte pour la première fois en 1957.
Elle se situe dans la membrane plasmique coté externe avec une activité
ATPasique du coté interne de la membrane côté cytosolique La Na + K + ATPase
pompe 3 Na + et le "chasse" hors de la cellule et fait rentrer 2K + dans la cellule
en consommant à chaque fois 1 ATP. Il y a une concentration plus élevée de potassium K+ à intérieur de la
cellule qu'à l'extérieur et inversement pour le sodium Na+. Pour maintenir son
intégrité la cellule doit pomper en permanence de sodium en le chassant vers
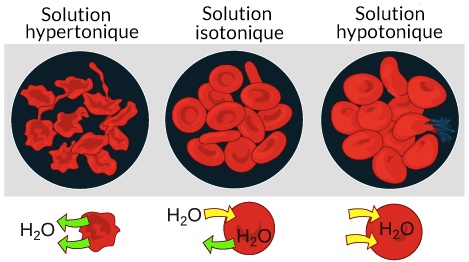
l'extérieur et inversement pour le potassium. Les phénomènes d'osmoses tendent à l'équilibrer la
concentration du sodium et du potassium entre l'intérieur et l'extérieur de la
cellule. Si la cellule devait être confrontée à une concentration de sodium
extérieur trop élevée (hypertonique), ce dernier aura tendance à rentrer dans le
cellule et la faire éclater car les mouvements de sodium sont suivis par des
mouvements d'eau (H20). Une solution hypotonique attirera le sodium vers
l'extérieur de la cellule et la fera se contracter jusqu'à la rupture membranaire.
En cas de concentration trop élevé de Sodium, le cellule va luter pour maintenir son intégrité en pompant vers l'extérieur le sodium qui sera entré par osmose. Mais si tous les canaux Na + K + ATPase sont tous saturés, le sodium continuera à rentrer suivi par l'eau, la cellule gonflera et explosera.
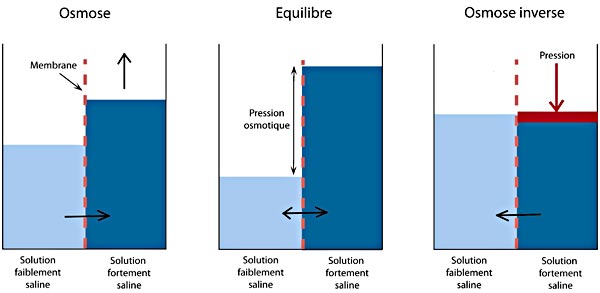
Expérience de la mise en évidence du principe de l'Osmose et la loi qui en dépend.
La loi de l'osmose ou de Loi de van 't Hoff est la suivante: π = RT C/M avec π qui désigne la pression osmotique en pascals (N/m²), R est la constante des gaz parfaits (8,31 J. mol-1. T est la température en Kemvin (°K), C = concentration pondérale du soluté, M = masse molaire.
1/ Technique d'analyse de la cellule
5/ Appareil de Golgi ou Dictyosome
13/ Complément sur la méthodologie de recherche en biologie cellulaire
![]()