Source Youtube: https://www.youtube.com/watch?v=n2cQP_260TM
Le noyau en division:
1/ La mitose :
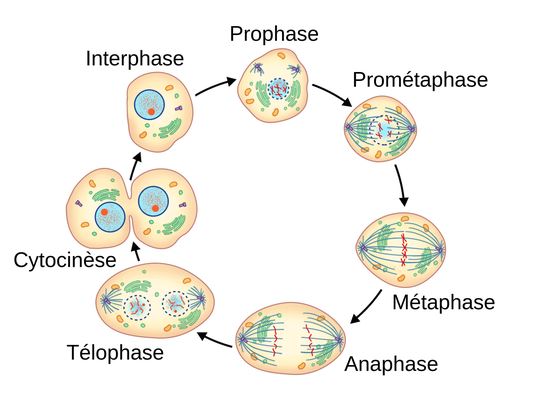
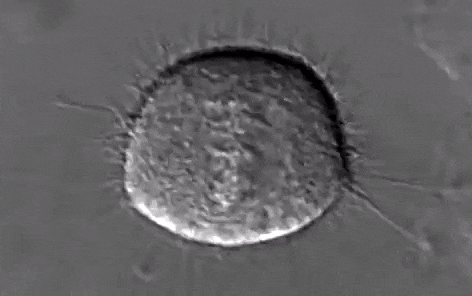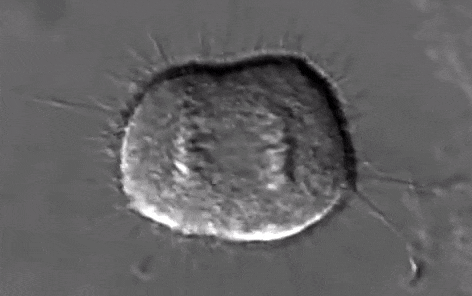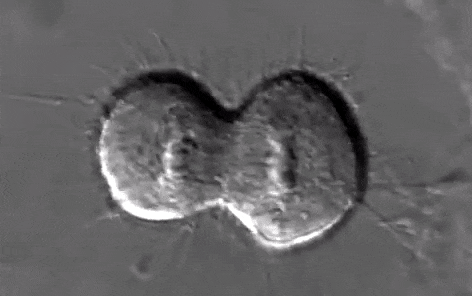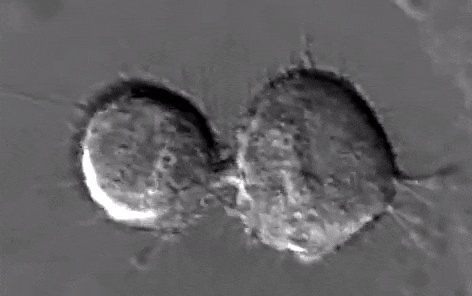
Schéma ci-dessus succinct de la mitose
Source Youtube:
https://www.youtube.com/watch?v=n2cQP_260TM
Elle pose deux problèmes :
- la compaction de l'ADN qui passe de l'état de chromatine à celui de chromosome et qui représente une compaction aux alentours de 5000 fois.
- la mécanique du tri des chromosomes, telle que chaque cellule fille reçoivent les mêmes chromosomes que la cellule mère.
C'est dans la phase G2 en général très courte que l'ADN commence à se compacter.
Étude de la spiralisation in vitro :
A partir de la chromatine isolée, en faisant varier la force ionique du milieu, on constate des modifications
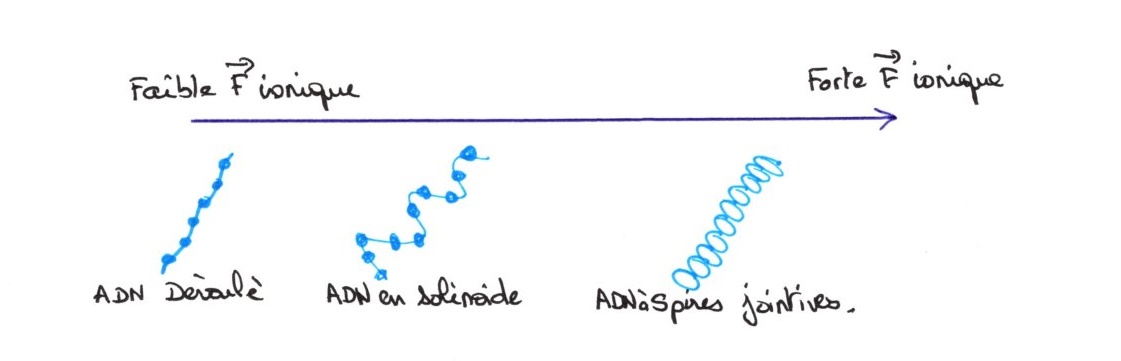
Dans un milieu de faible force ionique, on obtient une image artefactuelle car l'ADN est déroulé.
Puis, en augmentant la force ionique, on obtient une structure en solénoïde, la compaction est multipliée par 50. Ce phénomène est réversible si on diminue la force ionique. Lorsque les spires sont jointives, on obtient une compaction multipliée par 500. La force ionique fait varier les forces d'interactions hydrophobes entre les histones.
Il n'est pas certain que ce soit le même phénomène qui intervient in situ dans le modèle en solénoïde, deux histones H1 formeraient des interactions par leur partie hydrophobe ou bien, il y aurait formation de super nucléosomes.
On peut observer la formation de sous-unités de 6 à 8 nucléosomes.
a/ la prophase
C'est la phase la plus longue et la plus complexe. Elle se passe dans le noyau. En fin l'interphase, on voit pousser des microtubules, à partir des centrioles, en forme d'étoiles près de l'enveloppe nucléaire. Ce sont des microtubules astériens.
Le début de prophase est caractérisé par l'individuation des chromosomes en nombre fini. Ils comportent deux chromatides pour chacune, il y a un duplex d'ADN et en même temps mise en place du fuseau de division.
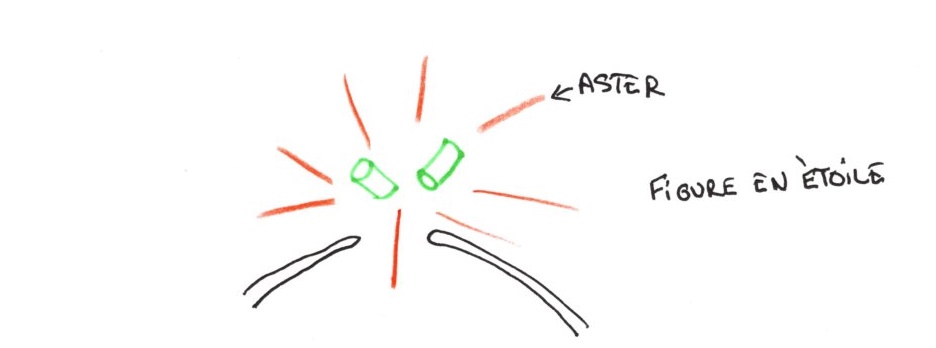
On observe la duplication des centrioles, avec formation d'un second diplosome. Chaque paire garde une orientation orthogonale.
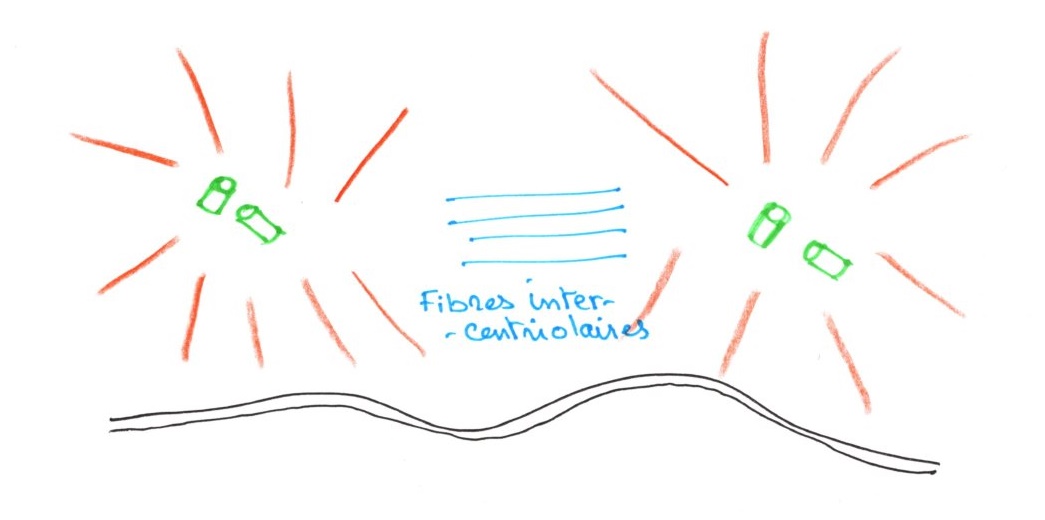
Entre les deux diplosomes, de nouveaux microtubules sont synthétisés. Leur allongement repousse les centrioles l'un de l'autre. On observe une dépression au niveau de l'enveloppe nucléaire.
En milieu de prophase, il y a migration des diplosomes vers les deux pôles de la cellule. Les microtubules intercentriolères se moulent autour de l'enveloppe nucléaire au cours de l'allongement.
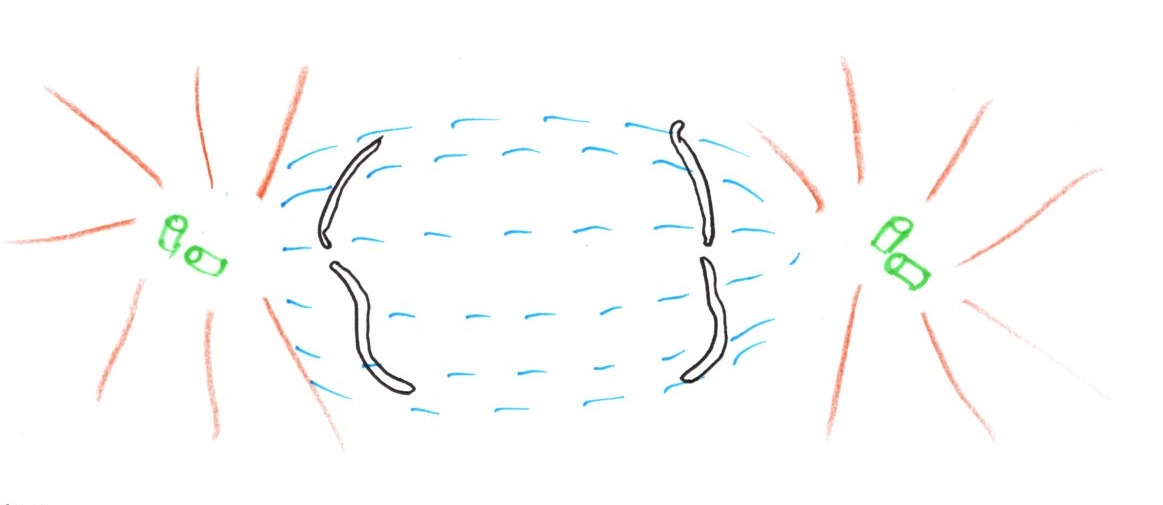
En fin de prophase ou prométaphase, l'enveloppe nucléaire se résorbe. Les microtubules polaires ne vont pas d'un centriole à l'autre car ils sont trop courts.
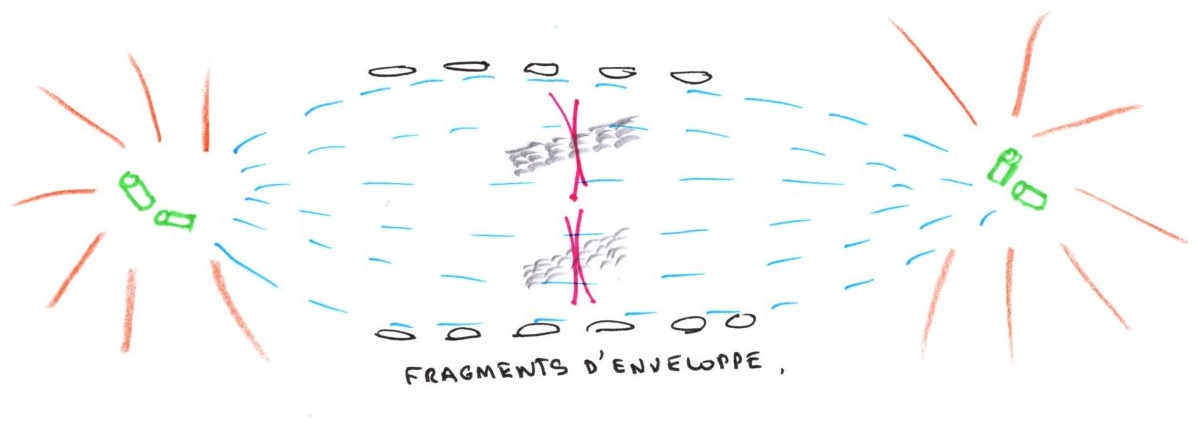
L'enveloppe nucléaire se fragmente et les fragments tendent à s'écarter. Le fuseau de division à la place du noyau est composé de microtubules polaires.
Les chromosomes sont libérés de l'enveloppe nucléaire, ils sont raccourcis, épaissis et flottent dans le nucléoplasme. On voit apparaître de nouveaux microtubules poussant au point d'attache les deux chromatides.

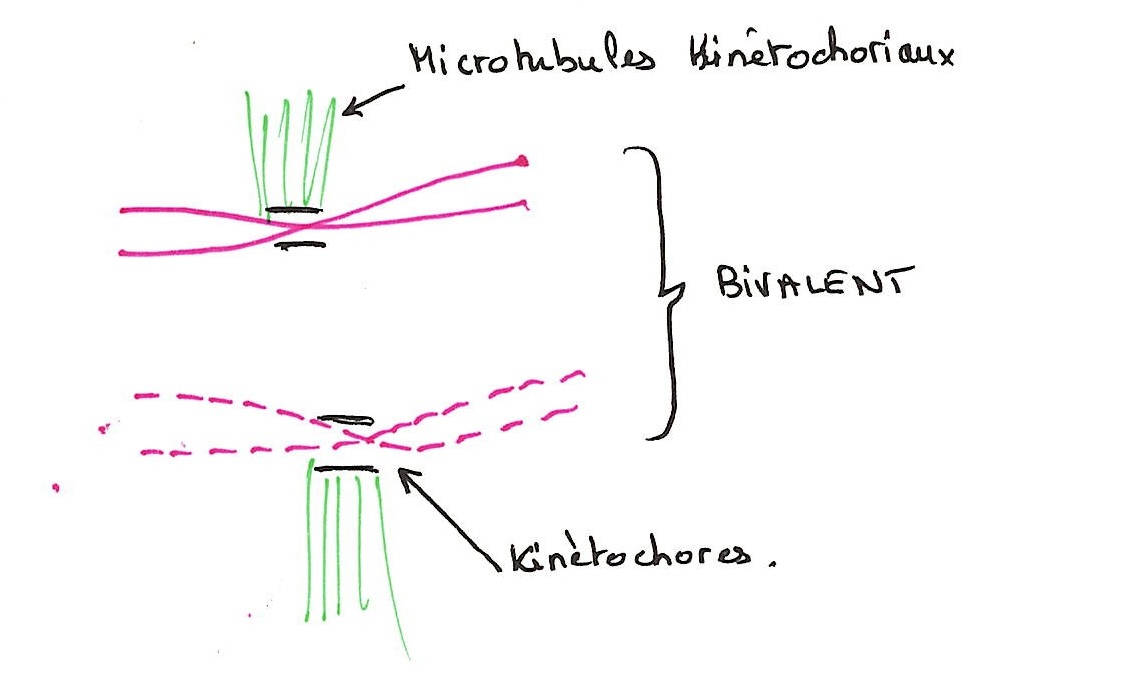
C'est à partir de la plaque kinétochorienne que se polymérisent les nouveaux microtubules kinétochoriens. Le kinétochore représente la zone de la plaque kinétochorienne.
b/La métaphase:
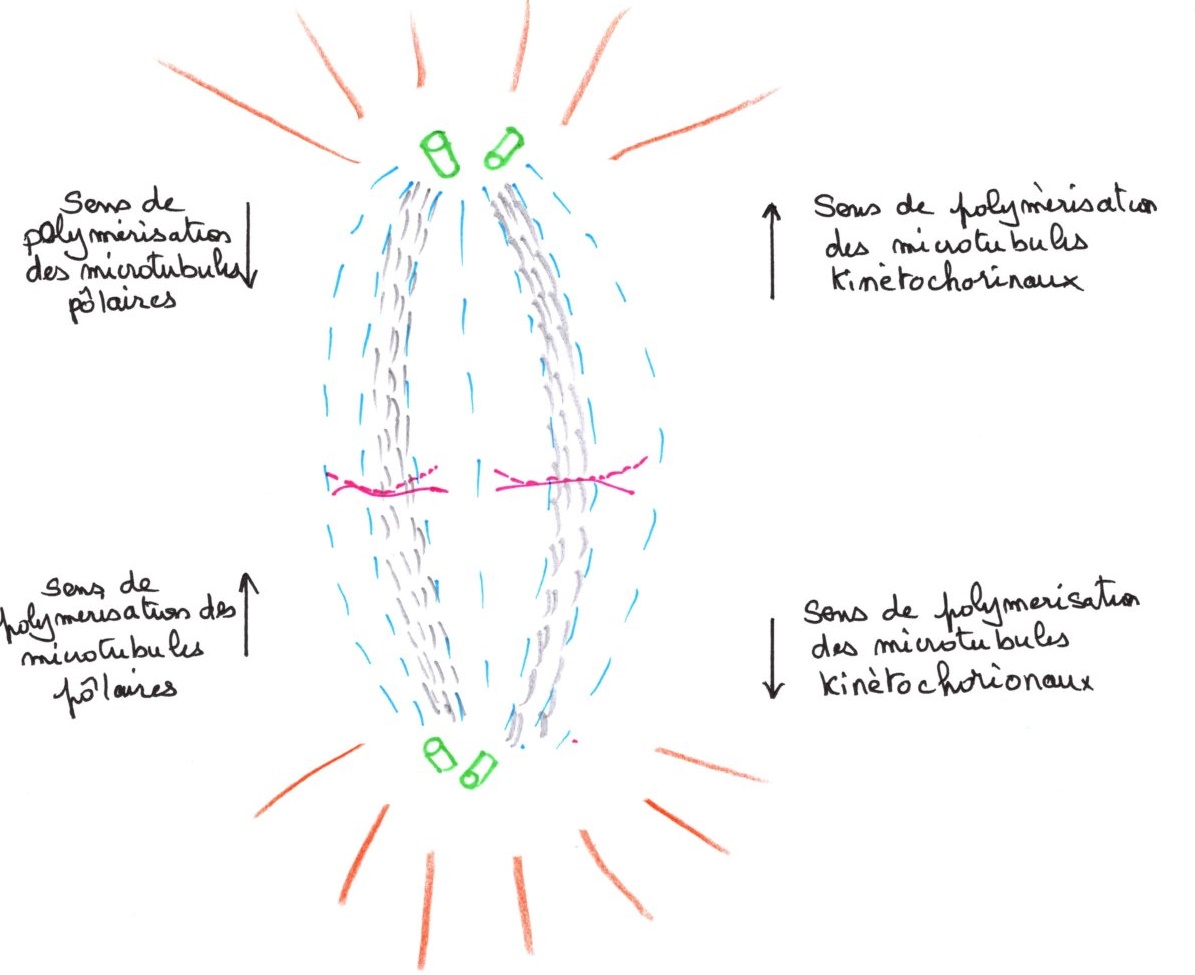
Il y a élongation des microtubules kinétochoriens. Leur orientation vers les pôles donne une figure de plaque équatoriale. Ces microtubules atteignent les deux pôles. Les chromosomes se répartissent sur un seul plan au milieu de la cellule. La plaque métaphasique ne se forme pas si on empêche l'élongation des microtubules kinétochoriens.
Il y a ainsi deux types de microtubules, les kinétochoriens et les polaires entre les pôles. Ils sont de même nature, mais d'origine différentes et parallèles entre eux.
C'est le stade le plus propice pour l'étude des chromosomes, car ils sont très condensés et étalés sur la plaque équatoriale.
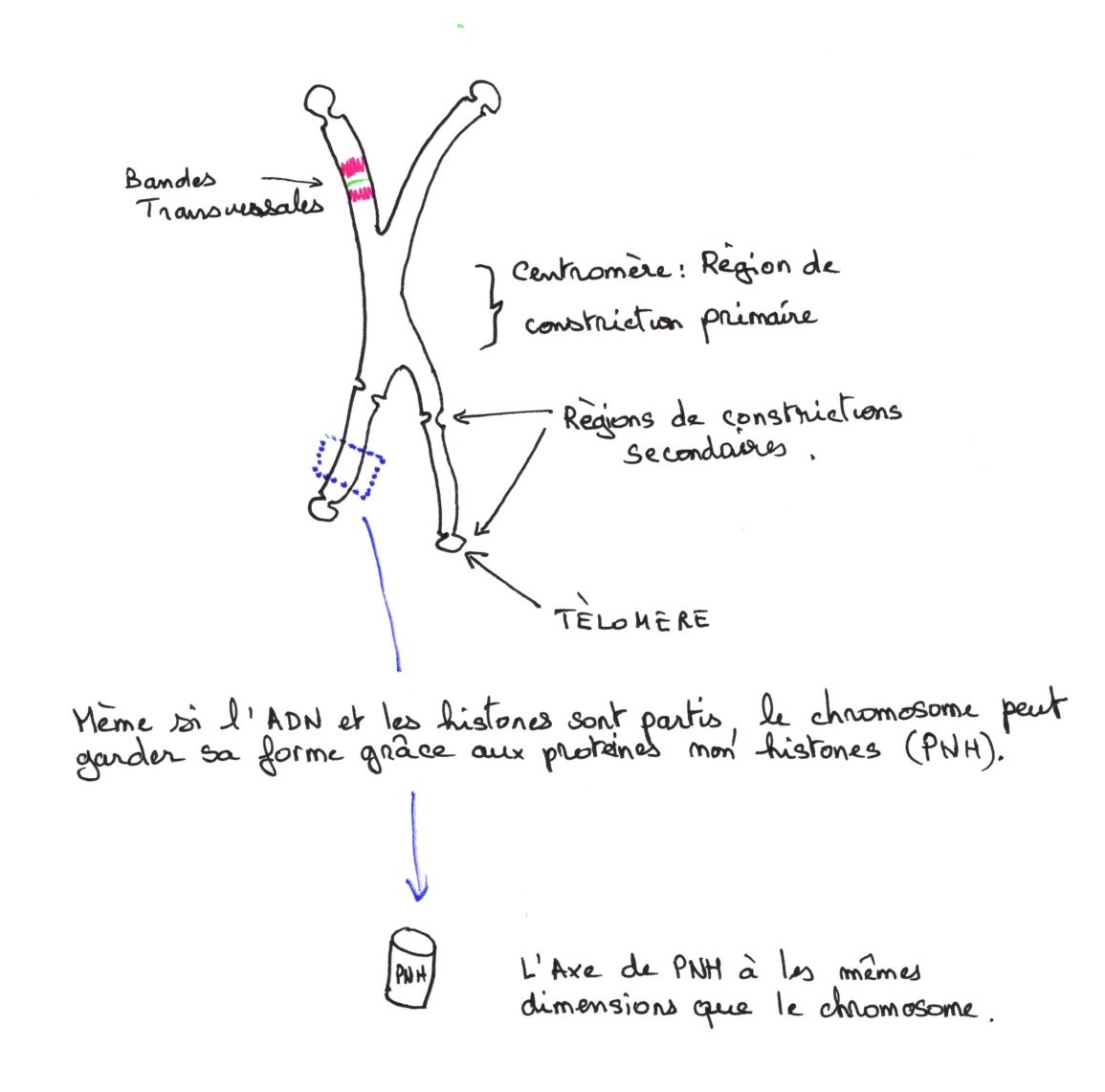
Le chromosome métaphasique :
Les deux chromatides sont strictement identiques. Le centre mère, région de construction primaire est l'endroit de fixation de la plaque kinétochorienne. Dans les régions de constructions secondaires, on trouve de l'hétérochromatine fortement compactée. L'ADN contenant le gène nucléaire est situé dans une construction secondaire invariable dans un type cellulaire donné.
Il y a des régions de différentes colorabilités:
- les constructions secondaires sont très colorables.
- le long des bras, des bandes plus claires sont toujours situées aux mêmes endroits et caractéristiques.
3 ordres de compaction :
- particules nucléosomique jusqu'au filament
- du filament au solénoïde
- du solénoïde au microconvules
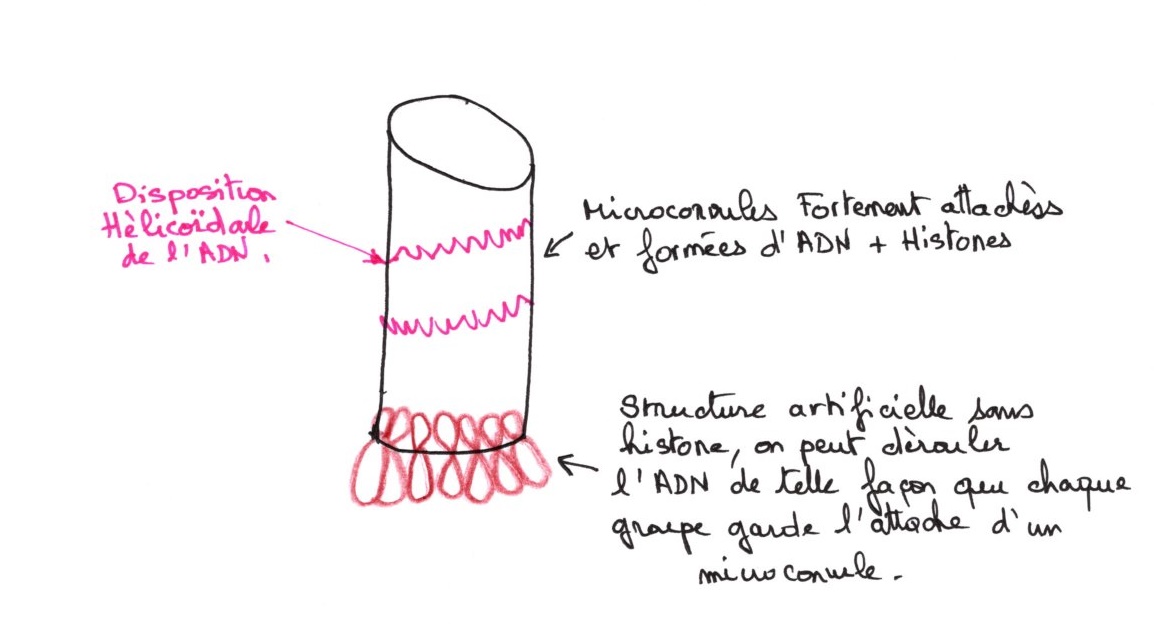
La carte morphologique du chromosome (son anatomie) est déterminée par:
- La longueur des bras est définie par la position du centromère.
- le nombre et la position des constrictions.
- le nombre et la position des bandes.
c/ L'anaphase :
Elle est caractérisée par l'ascension polaire des chromosomes, par un phénomène de dépolarisation des microtubules kinétochoriens qui se solubilisent dans la région des pôles et commence par séparer les paires de chromatides pour donner des chromosomes fils par clivage du centromère.
Il y a une ascension polaire des chromosomes. On peut voir au microscope électronique l'élongation du fuseau qui favorise la montée vers les pôles et la formation de l'interzone. Les microtubules polaires s'allongent, tandis que les microtubules kinétochoriens se raccourcissent.
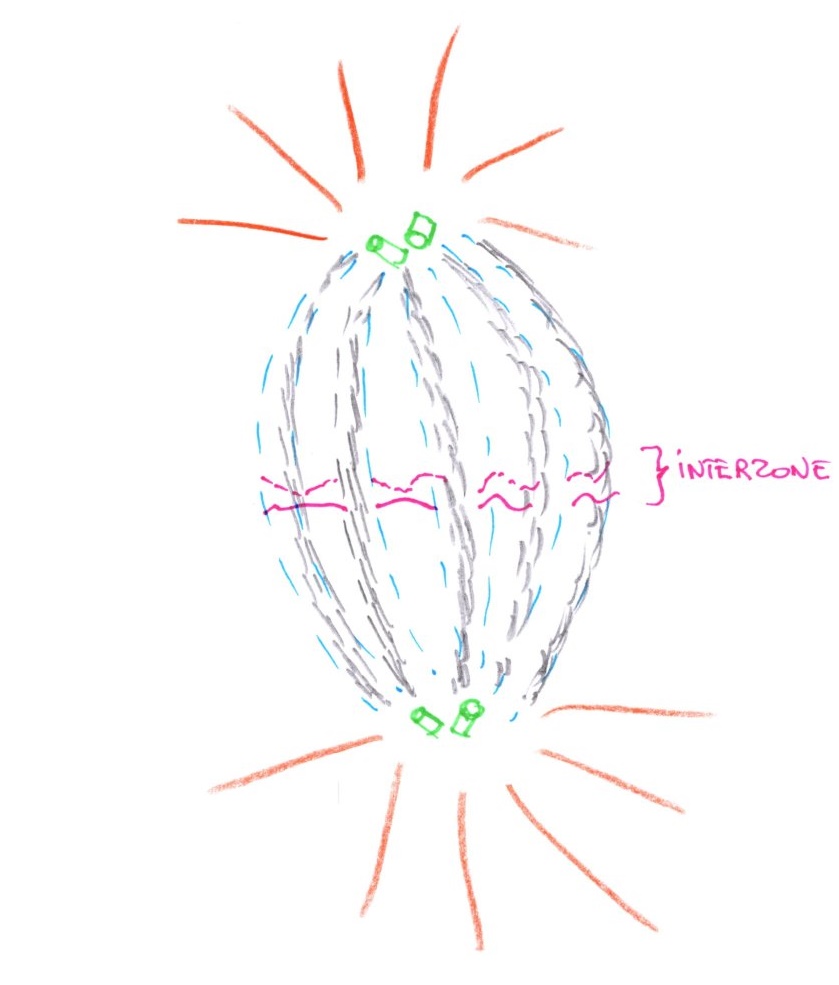
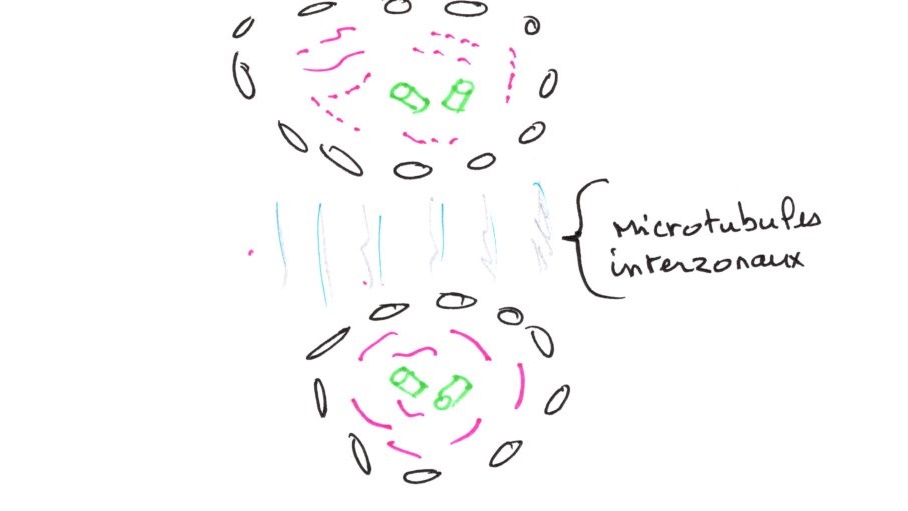
d/ La télophase :
Les deux lots de chromosomes se rassemblent autour des centrioles et commencent à se condenser. Les microtubules disparaissent. Des fragments d'enveloppes nucléaires se forment. Il y a condensation des microtubules dans l'Interzone. Les centrosomes sont plus près des pôles, les bras restants un peu en arrière du fait de leur inertie.
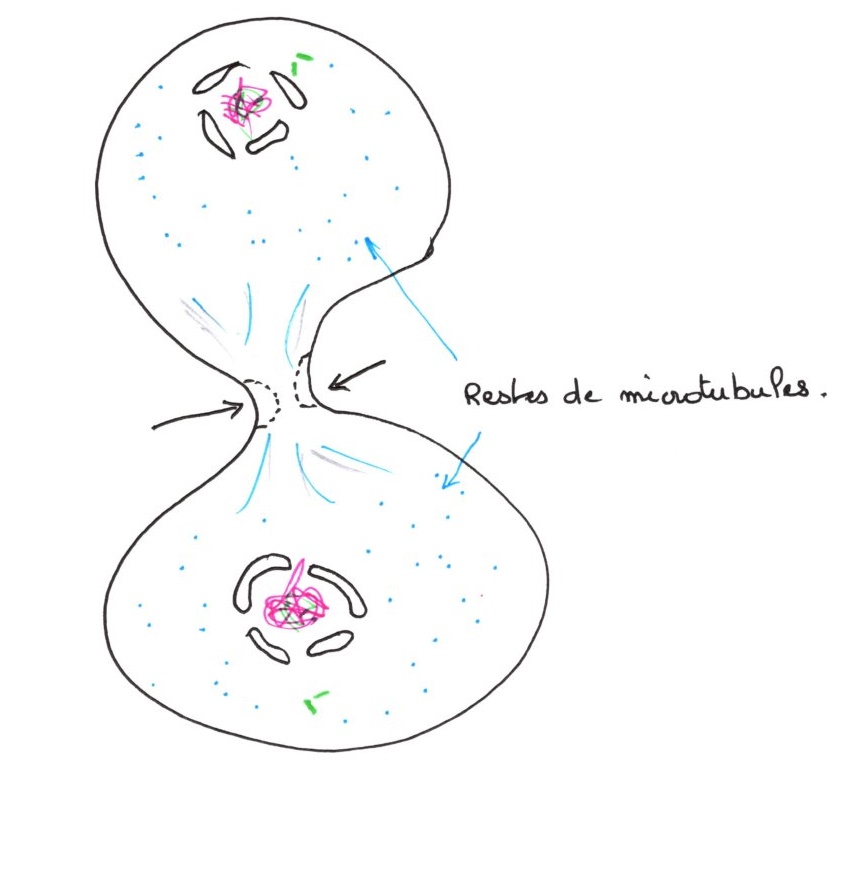
e/ La cytodiérèse :
C'est la séparation des cellules filles, les microtubules interzonaux se brisent au niveau de l'étranglement de la cellule qui progresse. Les restes de microtubules interzonaux fortement condensés, se dépolymérisent et permettent le rapprochement des deux étranglements. Il n'y a pas de changement du nombre de chromosomes, il s'agit d'une division équationnelle.
2/Ma méiose:
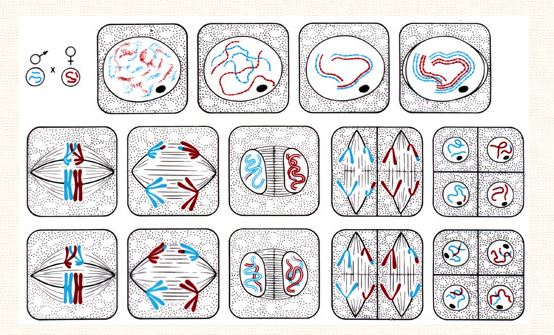
Elle concerne les cellules germinales (ovules et spermatozoïdes). Il s'agit des deux dernières divisions de ces cellules. Deux divisions successives avec une seule phase.
Source: https://www.youtube.com/watch?v=n2cQP_260TM
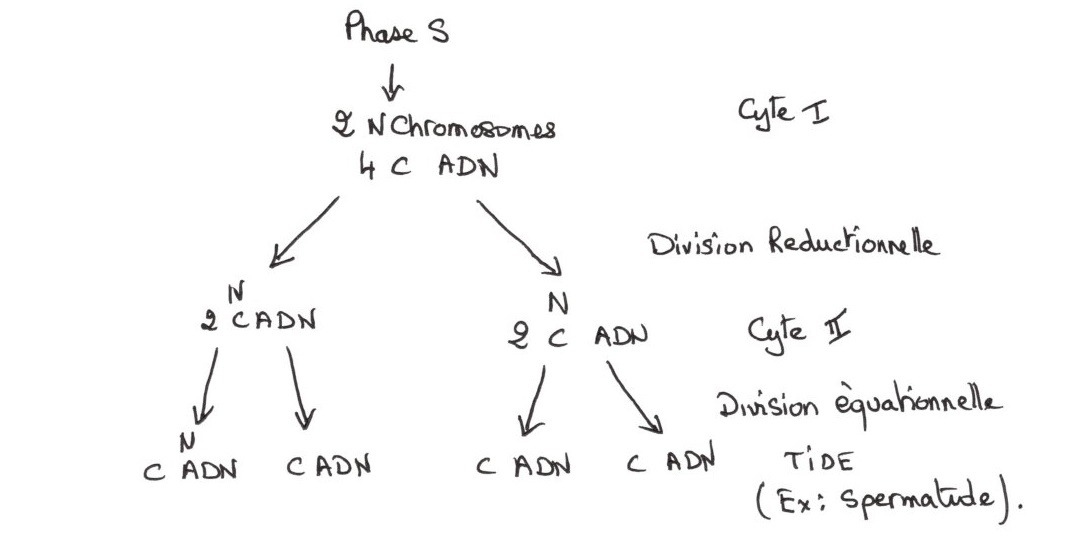
Cycle 1: A 2n chromosomes (stock: diploïdes), chaque chromosome est en double, issus de deux origines paternelles et maternelles.
Cycle2: Cellules filles, avec séparation des deux lots de chromosomes (stock haploïde), la quantité de départ est réduite de moitié.
TIDE: Les stock de chromosomique reste haploïde, la quantité d'ADN représente le quart de la quantité de départ. Le nombre de chromosome ne varie pas avec la quantité d'ADN.
Déroulement de la méiose:
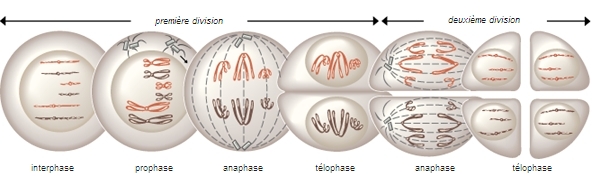
Prophase 1 puis métaphase 1, anaphase 1, télophase 1, cytodiérèse 1, puis, Prophase 2, métaphase 2, anaphase 2, télophase 2, cytodiérèse 2.
Souvent, la prophase 2 est court-circuité. Dans toutes ces phases, les mécanismes sont identiques à la mitose.
La prophase est divisée en 5 stades:
1:Stade leptotène:
Il y a apparition des chromosomes par un phénomène de compactage, mais ils sont encore longs et on ne distingue pas de constrictions secondaires. Les chromosomes homologues sont semblables car de même formes, mais ont une origine différente, ils ne sont pas appariés mais simplement proche les uns des autres.
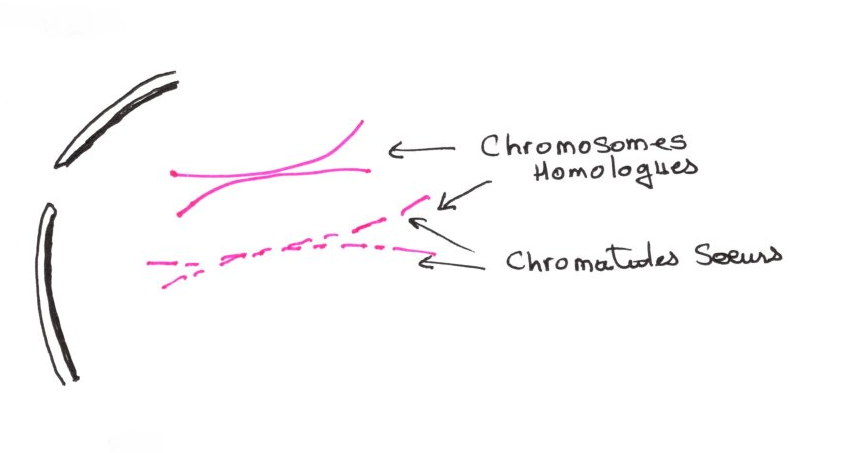
2:La stade zygotène:
A ce stade, il y a appariement des chromosomes homologues et leur fixation sur l'enveloppe nucléaire au niveau des télomères. Les deux chromosomes homologues se fixent au même endroit. Par raccourcissement et compactage des chromosomes, les centromères deviennent de plus en plus visibles. Dans la région des télomères où les chromosomes sont fixés sur la lamina, des PNH apparaissent dans un complexe d'appariement nommés SYNATONEMAUX. Ils se développent dans la région médiane et prolonge l'appariement.
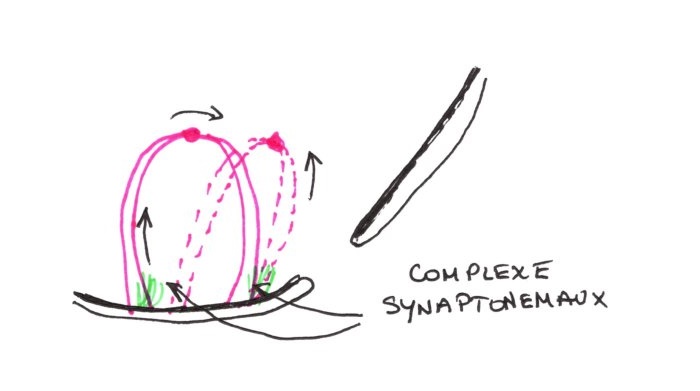
3:Stade Pachytène:
On ne distingue plus des chromatides entre elles, il y a
appariement étroit des chromosomes sur leur longueur par accroissement des
complexes. Les chromosomes se raccourcissent. Lorsque l'appariement se termine,
on ne distingue plus les deux chromosomes. On peut voir alors, l'ensemble en
nombre haploïde. Ce sont des bivalents. La forte condensation des bras de
chromosomes et la formations de bivalents favorisent les échanges de segments de
chromosomes. Si l'échange se fait entre les chromatides de deux chromosomes
différents, il y a une expression génétique différente. Ces échanges se font par
coupure et épissage grâce à des enzymes spécifiques.
Il est caractérisé par une
synapsis*
complète des paires de
synapsis* : Le synapsis est nommé aussi appariement, c'est l'union des chromosomes homologues qui donnent des tétrades (des groupes de 4 chromatides).

4:Stade diplotène:
On observe:
-Les chiasmes deviennent visibles.
-Les centromères s'éloignent.
-Les télomères se séparent de l'enveloppe nucléaire.
-Les bivalents sont libres, tandis que les chromosomes homologues sont liés.

5:Diacinèse:
Il y a éloignement des centromères, les chiasmas glissent vers les extrémités des chromatides.
Il y a mouvement divergent des centromères. Le reste de l'appareil mitotique se met en place. On observe la résorption de l'enveloppe nucléaire en prémétaphase.

La métaphase 1
L'enveloppe nucléaire se résorbe. Une plaque kinétochoriale se forme au niveau des centromères. Il y a élongation des microtubules kinétochoriaux. Ceux d'un même chromosome vont vers un même pole. Les chromosomes se placent dans le plan médian de la cellule pour former une double plaque équatoriale.
L'anaphase 1:
Les chromosomes "entiers" montent aux deux pôles et à chaque pole, on trouve la moitié de la totalité des chromosomes. Il y a ségrégation aléatoire des chromosomes paternels et maternels. Ce brassage des chromosomes entraîne des conséquences génétiques. C'est un brassage interchromosomique. Il se rajoute au brassage intrachromatique dû au CROSSING-OVER.
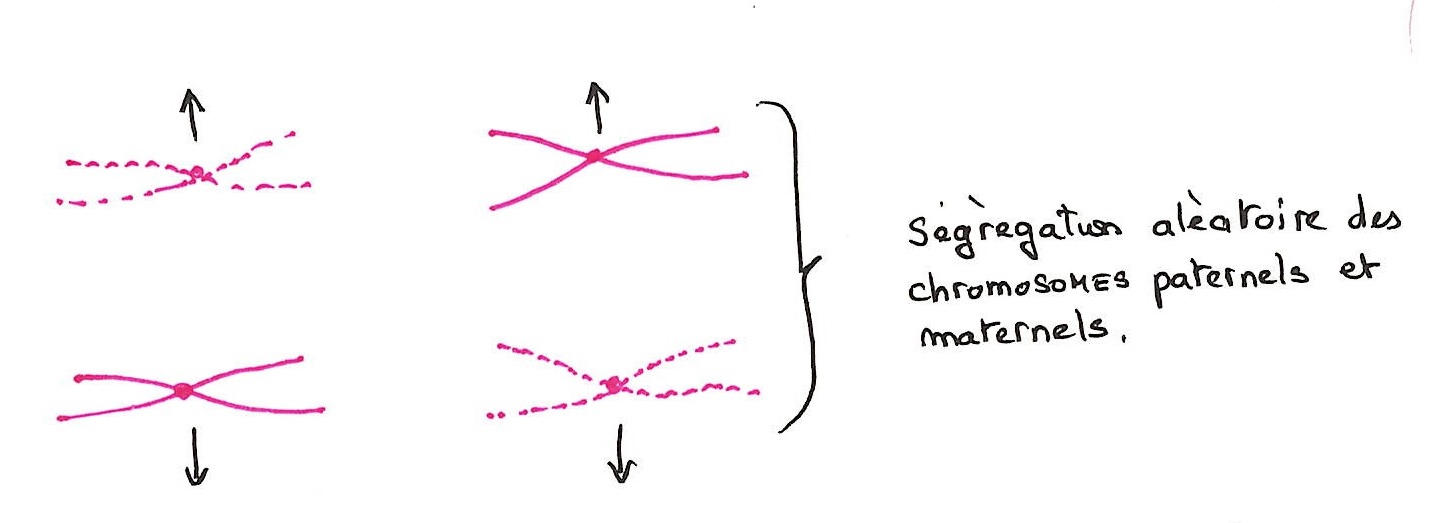
En télophase 1, les chromosomes se décondensent et l'enveloppe nucléaire se réforme.
La Deuxième division de méiose se passe comme une mitose habituelle. C'est une division de type équationnelle se déroulant à l'état haploïde. Après des deux divisions, chacune des 4 chromatides est dans une cellule fille différente. Les quatre cellule filles sont donc toutes différentes.
1/ Technique d'analyse de la cellule
5/ Appareil de Golgi ou Dictyosome
13/ Complément sur la méthodologie de recherche en biologie cellulaire
![]()